Навигация
Епідеміологічне значення окремих видів і штамів малярійних паразитів
114995
знаков
6
таблиц
6
изображений
1.2.1 Епідеміологічне значення окремих видів і штамів малярійних паразитів
Людина може захворіти на малярію внаслідок природної спорозоїтної інфекції після укусу зараженого комара або штучної трофозоїтної інфекції при введенні здоровій людині певної кількості крові, яка містить шизонти. Епідеміологічне значення має тільки природне зараження через комара, тому що Інфекція при цьому характеризується більш; тривалим перебігом, більшою частотою та упертістю повторних рецидивів. Крім того, штучні зараження бувають рідко при терапевтичній малярії або при випадковій трансфузії крові донора, яка містить шизонти. До речі, останнім часом описано багато випадків зараження малярією при переливанні крові від донорів—латентних носіїв, тому обслідування всіх донорів є обов'язковим з метою виключення латентних носіїв.
Кожний рецидив малярії, викликаний поширеним розмноженням шизонтів у периферичній крові, має велике епідеміологічне значення, тому що при всякій шизогонії утворюються також гаметоцити, які дають можливість зараження комарів і дальшого поширення інфекції.
Однак слід пам'ятати, що не кожне збільшення шизогонії у периферичній крові у всіх випадках супроводжується рецидивом з характерними гарячковими приступами. Часто у носіїв, особливо в епідемічний сезон, при частковому ослабленні супровідного імунітету, кількість паразитів може значно збільшитись, але не досягати „пірогеннрго порогу", тобто тієї кількості паразитів, при якій у даного хворого почнуться клінічні вияви малярії; в таких випадках бувають так звані „холодні рецидиви", тобто прояви посиленої шизогонії крові без гарячкових приступів.
Іноді в крові виживають найбільш стійкі форми паразитів — гаметоцити; в цих випадках шизонти важко виявляються в крові, і спостерігається явище чистого гаметоцитоносіння. Ці холодні рецидиви і гаметоцитоносіння мають величезне значення в епідеміології малярії, тому що, не супроводжуючись видимими виявами хвороби, вони залишаються непомітними і, не будучи правильно ліковані, є постійним джерелом зараження комарів і дальшого поширення малярії.
Гаметоцитоутворення є важливим біологічним явищем у розвитку і розмноженні паразита, яке забезпечує йому збереження його виду в природі, тому що гаметоцити здатні до дальшого розвитку в шлунку комара, а шизонти гинуть; крім того, гаметоцити пристосовані до більш тривалого виживання в крові людини і більш резистентні щодо хініну та акрихіну/ а також імунної захисної реакції організму.
Наступним дуже важливим моментом для епідеміології малярії є те, що плазмодії в більшості випадків схильні відносно довго залишатись в організмі зараженої людини, в середньому 2—3 роки, причому паразитовосіння має циклічний характер: чергуються періоди посиленої життєдіяльності паразита, посиленого розмноження його у периферичній крові, Що супроводжується клінічно виявленими або безсимптомними холодними рецидивами, або настають латентні періоди, коли інфекція не виявляється в крові, і не має ніяких зовнішніх виявів хвороби, після чого іноді цілком несподівано можуть настати нові серії рецидивів.
Оскільки нині немає ніяких способів для з'ясування питання про те, чи триває у людини латентне носіння паразитів і чи можуть ще настати повторні рецидиви, з епідеміологічної точки зору кожного захворілого на малярію треба розглядати як потенціального паразитоносія і тримати його під спостереженням протягом середнього строку тривалості інфекції малярії, тобто коло двох років.
В епідеміології малярії мають також значення сезонні посилення активності паразитів різних видів і штамів, що виявляється здатністю давати більш часті періоди посилення периферичної шизогонії, що часто супроводжується приступами в певні сезони року, характерні для кожного виду. Найбільша активність Pl. vivax припадає на весняний період і початок літа, залежно від географічного положення місцевості, І збігається з періодом встановлення середньої декадної температури 16 — 20°, що звичайно відповідає виплоджуванню першої і початку другої генерації анофелес. Найбільша активність Pl. falciparum відповідає періодові більш високих літніх температур від 22° і вище, що для більшості південних областей СРСР буває в другу декаду липня і збігається з максимальною скупченістю анофелес. Та обставина, що максимальна сезонна активність Рl. vivax і Pl. falciparam збігається з періодами активності комарів, обумовлює те, що обидва ці види паразитів при певних умовах можуть давати епідемічні спалахи, причому навіть у тих місцевостях, де поширення цього виду зустрічає перешкоду. Тому Pl. malariae, для якого максимальне підвищення сезонної активності припадає на останні місяці літа і перші осінні місяці, коли активність комарів уже знижується у зв'язку з підготовкою до переходу на зимівлі, ніде на території СРСР не давав епідемічних спалахів.
Вищесказане робить зрозумілим важливість щомісячного обліку первинної реєстрації малярійної захворюваності і складання помісячних кривих. Аналіз цих кривих дає цінні дані: наявність весняно - літнього підвищення, так званий „північний тип” кривої, говорить за превалювання поширення терціани, осіннє підвищення, або „південний тип” кривої, характерний для місцевостей з поширенням тропічної малярії; з другого боку, високе осіннє підвищення помісячної кривої захворюваності для місцевостей, де переважно поширена тільки терціана, вказує на ріст свіжих захворювань цього року і дає несприятливий прогноз для наступного року.
Епідеміологічні особливості малярії в значній мірі залежать від видової належності збудників, тому нижче . наводяться основні відомості для характеристики кожного виду.
1. Pl. falciparum Welch. 1887, — вид, що дає в межах СРСР ряд значних ендемічних вогнищ, розміщених у місцевостях з більш жарким кліматом (Середня Азія, Закавказзя, Північний Кавказ, Нижнє Поволжя), а також, що періодично буває причиною серйозних епідемічних спалахів у місцевостях з помірний кліматом, зокрема на півдні України. Цей вид паразита викликає захворювання найбільш тяжкі щодо своїх клінічних проявів, причому вже перші приступи можуть закінчитися летально, але разом з тим ця форма щодо-своєї тривалості є найкоротшою: середня тривалість паразитоносіння, очевидно, дорівнює 12 місяцям. Якушева вважає, що крайній строк настання клінічно виявлених рецидивів становить 13 місяців, а холодних рецидивів — 17 місяців. Велика кількість спостережень тривалості паразитоносіння при тропічній малярії показала, що, приблизно в одній третині випадків, інфекція Pl. falciparam після первинної атаки приступів і серії ранніх рецидивів дуже рано може закінчитися цілковитим видужанням; в інших випадках можуть бути повторні рецидиви, а також повторні повернення гаметоцитоносіння протягом усієї зими і ранньої весни: в більш пізні весняні місяці паразитоносіння Pl. falciparum витискається носінням Pl. vivax.
В кінці літа наступного року після зараження можуть знову настати клінічні рецидиви, після чого Звичайно настає гаметоцитоносіння, яке може тривати від 7—10 днів до 1 місяця; гаметоцити є найбільш стійкими до хініну і імунними до захисних реакцій організму. Супровідний імунітет утворюється важко і він дуже лабільний, тому можливі повторні інокуляції паразитів того ж виду і штаму, що призводить до частих повторних клінічно виявлених захворювань, з наступними періодами гаметоцитоносіння.
В зв'язку з вище переліченими особливостями Pl. falciparum треба вважати найбільш небезпечним видом в епідеміологічному значенні. При певних кліматичних умовах він може давати виявлені ендемічні вогнища, в яких завжди можливі окремі, дуже сильні епідемічні спалахи, іноді з високою смертністю.
2. Pl. vivax Grassі et Felettі, 1890, — малярійний паразит більше поширений у місцевостях з помірним кліматом, у південних вогнищах з превалюванням Pl. falciparum зустрічається в невеликих кількостях і майже зовсім відсутній на тропіках. Тривалість паразитоносіння вдвоє більша, ніж при тропічній малярії, в середньому — близько 2 років, але звичайно не перевищує 3-х років. За нашими спостереженнями, у роки загального зниження малярійної захворюваності тривалість паразитоносіння зменшується і в ряді випадків хвороба може закінчитися раннім видужанням, скороченням кількості рецидивів, що настають наступної весни: взагалі паразитоносіння характеризується значною кількістю рецидивів як ранніх, такі віддалених.
Ця схильність до частих рецидивів значно збільшується в роки епідемічних підвищень захворюваності на триденну малярію. Випадки холодного паразитоносіння і гаметоцитоносіння досить часто зустрічаються, але тривалість їх значно менша, ніж при інфекціях Pl. falciparum; гаметоцити теж менш стійкі. Тривалість шизогонії у Pl. vivax на 12 годин більша, ніж у Pl. falciparum, тому нагромадження паразитів у крові відбувається більш повільно, у зв'язку з чим і тривалість інкубаційного періоду трохи більша, тобто коло 15 діб. Проте в більш північних місцевостях Pl. vivax дає інфекції з тривалою інкубацією до 6 — 11 місяців, тому в осіб, що заразилися в кінці літа, перші вияви захворювання спостерігаються весною наступного року. Можливість цієї тривалої інкубації має певне "епідеміологічне значення і пояснює масову появу свіжих захворювань на триденну малярію на початку нового епідемічного сезону ще до початку періоду активності комарів.
Можливість тривалого інкубаційного періоду є одним з проявів біологічної пристосованості паразита для збереження свого виду в природі. Тому що в північних областях з більш низькими середньодобовими температурами передавання інфекції протягом того епідемічного сезону не завжди буває можливим завдяки короткому літу і ранньому відльоту комарів на зимівлю, то й активність паразита та активність переносника поступово пристосувались до того лі самого часу.
Завдяки вищевказаним біологічним особливостям гі, vivax при певних кліматичних умовах, у місцевостях з помірним кліматом можуть створюватись серйозні ендемічні вогнища триденної малярії; в окремі роки можуть виникати величезні епідемічні спалахи, які, проте, по інтенсивності наростання захворюваності і по тяжкості клінічних виявів поступаються перед іронічною малярією.
3. Pl. malariae Laveran, 1881 — вид паразита, що обумовлює захворювання в період осені і початку зими, причому паразитоносіння частіше виявляється в наших кліматичних умовах у період майже повного припинення активності комарів. Це, очевидно, є основною причиною, яка обмежує поширення цього виду, і захворювання на чотириденну малярію зустрічається майже скрізь, але у вигляді поодиноких спорідичних випадків. Тривалість паразитоносіння ще точно не встановлена, але є підстава гадати, що вона найбільш тривалою, в середньому — до 3 років. Цикл шизогонії найбільш тривалий (72 години), материнська клітина дрібниться тільки на 8 10 мерозоїтів, тому численне зростання паразитів при шизогонії відбуваюся найбільш повільно, в зв'язку з чим інкубаційний період при інфекціях цим видом паразита найбільш тривалий — до 21 дня. Для чотириденної малярії характерні повторні окремі рецидиви з наступним при палим паразитоносінням.
Штами малярійних паразитів. Останнім часом, крім давно відомих даних про види малярійних паразитів (Рl. vivax, Pl. malariae, Pl. falciparum), що викликають різні форми малярії, особливий інтерес становлять цілком нові дані про штами або раси, які можна констатувати серед плазмодій одного виду. Ці штами можуть бути розрізнювані тільки за своїми біологічними властивостями; ступенем вірулентності, ступенем заразливості для людини і комарів, ступенем резистентності до діяння протималярійних медикаментів та імунологічними особливостями.
Нині не виявлено ніяких морфологічних ознак, які давали б можливість розрізняти окремі штами, але не виключена ймовірність того, що в майбутньому при поліпшенні методів забарвлення і мікроскопічної техніки будуть виявлені і морфологічні особливості, окремих штамів (Coatney, Joung). Вже давно при вивченні, італійських та індійських штамів Pl. falciparum вдалося помітити, що перші з великою постійністю дають більш тяжкий клінічний перебіг, більшу тривалість первинною гарячкового періоду, більшу стійкість до діяння хініну і необхідність великих доз його для припинення гарячки. Індійські штами давали коротше і легше захворювання, що легко купірується хініном. Аналогічні дані наводить Kortweg для Голландії — для привезених мадагаскарських і для голландських штамів.
Багато цінного для вивчення штамів дали терапевтичні перещеплювання малярії (для лікування паралітиків): окремі штами протягом тривалого строку прищеплювались на тисячах хворих і незважаючи на це, стійко зберігали свої особливості, тобто схильність давати легкий або тяжкий перебіг, частоту рецидивів та інтенсивність гаметоцитоутворення. Практично дуже важливим є те, що імунітет і несприйнятливість до, нових інокуляцій цим самим штамом, що розвинулися після повторних заражень, є суворо специфічними для даного штаму, тому що інфікування новим штамом, привезеним з інших місцевостей, дає гостре захворювання у цих імунних осіб.
У нас для Pl. vivax можна констатувати також північні і південні штами. Північні штами дають довгий інкубаційний період (до 8 — 11 місяців), південні— короткий інкубаційний період (коло 10 днів).Ця схильність давати короткий або довгий інкубаційний період стійко зберігається у певного штаму, незважаючи на повторні перещеплювання паралітикам. Крім того, штами відрізняються також іншими ознаками — тяжкістю перебігу, кількістю паразитів у периферичній крові, інтенсивністю гаметоутворення, стійкістю до акрихіну.
Ці нові дані про штами в окремих видів плазмодій є дуже цінними для епідеміології і дають можливість детальнішого тлумачення багатьох явищ, зокрема причин епідемічних спалахів триденної малярії: завезення в райони, навіть з великим поширенням місцевої малярії, може бути однією з істотних причин спалаху, тому що населення, яке мало деякий імунітет до місцевих штамів, може виявитися неімунним до завезених штамів.
1.2.2 Зараження комарів, спорогоніяНе кожна самиця анофелес, що нассалася крові і аметоцитовосіи, заражається, не кожне зараження, яке настало, доходить до стадії утворення спорозоїтів. Нарешті, не завжди спорозоїти, що перебувають у слинних залозах, можуть викликати нове зараження людини, тому що при певних умовах спорозоїти втрачають свою інфекційність. Гаметоцити, що попали н шлунок комара разом з кров'ю, перетворюються на гамети, відбувається копуляція чоловічих і жіночих гамет і утворюється зигота. Ці початкові етапи статевого розмноження паразитів у тілі комара потребують температури не менше 16° для Pl. vivax, 17 — 18° для Pl. falciparum. Крім того, щоб відбулося зараження комара, потрібна кількість в 1 мм3 крові паразитоносіння 10 гаметоцитів чоловічих і жіночих, з переважанням останніх.
Однак, як показав Вещезоров, немає прямої залежності між зараженням комара і кількістю гаметоцитів у крові малярика і іноді зараження комара не вдавалося, хоч в крові хворого було 348 гаметоцитів в 1 мм3. Ще Джеме (James) вказував, що при штучному зараженні вдається заразити не всіх комарів. Нарешті, щоб відбулося зараження комара від гаме- і тоцитоносія, останній не повинен одержувати плазмоциду, приймання якого різко зменшує можливість зараження комара.
Тимчасове зниження температури нижче 16°, що збіглося з тим часом, коли ооцисти ще не встигли і утворитися під слизовою шлунка комара, може призвести до загибелі зигот і припинення процесу спорогонії, коли ж ооцисти вже утворилися, то тимчасове ; зниження температури тільки затримує їх розвиток, але не призводить до їх знищення. Це має велике практичне значення для епідеміології малярії, тому що показує, що тимчасові зниження температури, які можуть бути влітку, не завжди припиняють спорогонію, а тільки затягають строки її закінчення. Тільки тривалі зниження температури в осінньо-зимовий період, коли комарі перебувають на зимівлях, приводить до генерації ооцист.
Тривалість всього процесу спорогонії до останнього; її етапу, тобто до нагромадження спорозоїтів у слинних залозах, при 16 — 17° дорівнюють для Pl. vivax 53 дням; це приблизно відповідає строку тривалості життя літньої самиці An. maculipennis; тому в місцевостях з такою середньою літньою температурою, більше самиць гине раніше, ніж вони зможуть передав вати малярію, тому в цих місцевостях малярія aбo зовсім не зустрічається або буває у вигляді поодиноких, спорадичних випадків.
При підвищенні середньодобової температури тривалість спорогонії зменшується і при 30° буде тільки 7 днів; при оптимальній температурі для виживання комарів, при 25 — 26°, тривалість спорогонії дорівнює 10 — 11 дням. Для Pl. falciparum, щоб почався процес спорогонії, температура повинна бути не нижче 17 — 18°. Залежність початку і тривалості спорогонії від температури робить зрозумілим, чому в місцевостях з більш жарким літом процент заражених комарів вищий, отже, може бути вищою і малярійна захворюваність; зрозуміло також, що в місцевостях з помірним кліматом в окремі роки з більш високою літньою температурою можна чекати збільшення процента заражених комарів, отже й збільшення захворюваності на малярію.
Отже оптимальна, висока температура у літні місяці обумовлює високу малярійну захворюваність внаслідок того, що: 1) при цьому скорочується тривалість спорогонії, 2) при наявності оптимальної вологості збільшується тривалість життя окриленого комара, нарешті, 3) при більш високих температурах прискорюється процес перетравлювання крові в шлунку комара і відкладання яєць, а разом з цим з'являється значно частіша потреба нападати на здобич і ссати кров повторно. Але, разом з тим спорозоїти можуть дуже довго зберігатися в слинних залозах у комара — до 3 — 4 місяців у зимуючої самиці і до природної! смерті у літньої самиці; тому чим частіше заражена самиця повторно нападає на людей, тим більша кількість нових заражень може відбутися при цьому. Однак, хоч спорозоїти можуть зберігатись довгий час у слинних залозах комара, через деякий час, як показав Барбер (Barber), вони втрачають здатність заражати ; людину, незважаючи на те, що залишаються живими. В слинних залозах у зимуючої самиці такі зміни відбуваються через 1 — 11/2 місяці. Тому самиці, які вилетіли весною після зимівлі, вже не можуть весною заражати нових людей.
Самиці, що вилетіли після зимівлі, можуть знову заражатися, але практично в наших кліматичних умовах більша частина самиць, що перезимували, гине раніше і не доживає до періоду із стійкою середньодобовою температурою 16°, при якій тільки можливий початок процесу спорогонії. Тому в місцевостях з помірним кліматом самиці, що перезимували, відіграють дуже невелику роль у виникненні нових заражень людей.
Спостереження над заражуваністю різних видів і анофелес у різних кліматичних умовах показали неоднакове ставлення до зараження тим чи іншим видом плазмодія; проте епідеміологічне значення різних видів анофелес буде неоднаковим у різних місцевостях і залежить від екологічних умов, в яких перебуває комар і в тих чи інших місцевостях.
1.2.3 Епідеміологічна характеристика деяких моментів біології та екології малярійного комараНе знаючи біології та екології малярійного комара, не можна дати оцінку багатьом епідеміологічним явищам і правильно організувати протималярійні заходи. І
Нижче наводиться тільки частина відомостей, які мають практичне значення для розуміння деяких питань епідеміології малярії. Більш докладний виклад і їх теоретичне обґрунтування подає Беклемішев.
Практично являють собою інтерес:
1. Розподіл анофелес у природі і ступінь «домашності» його.
2. Умови, що визначають шукання і напади комара і а здобич.
3. Тривалість життя літньої самиці і співвідношення між її віком і зараженістю.
4. Зміна поведінки комара в період переходу до зимівлі та епідеміологічне значення цього явища.
5. Епідеміологічне значення підвидів.
Для малярійного комара потрібні певні мікрокліматичні умови, з яких на першому місці слід поставити відносну вологість повітря. Ця вологість у сполученні з певною температурою є обов'язковою умовою дія підтримування водного балансу комара. Крім того, па днюваннях комарові сприяє відсутність протягів та ослаблене освітлення. Ці умови характеризують днювання комара; вони значною мірою залежать також від добових і сезонних кліматичних змін, які бувають різними для кожної місцевості.
Не менш важливі дані фауністичного характеру, які зазначають видовий і підвидовий стан комариної популяції, тому що біологія комара значною мірою залежить від його видової належності. В межах того самого виду нерідко спостерігаються коливання, які залежать від місцевих кліматичних умов, наприклад, поведінка самиці An. m. messeae на території Середнього Уралу (Магнитогорськ) визначається відносною вологістю не менше 65°, в той час, як в Туркменії (Каратала) активність самиці пробуджується при значно нижчій вологості (45—50°/0).
Денні сховища комара, звичайно, не можуть бути однаковими; комар віддає перевагу тим з них, які є більш сприятливими в екологічному відношенні. Тому слід розрізняти типові і випадкові днювання: в останніх комар залишається недовго.
З епідеміологічної точки зору являє значний інтерес питання, наскільки житло людини є місцем денного перебування комара. Житла людини можуть бути цілком сприятливими умовами для денного перебування комара тільки в тих випадках, коли вони найбільше підходять до природних умов, біологічно необхідних для комара. В противному разі вони є тимчасовими і «вимушеними». Весною і восени житла людини більше наближаються до природних умов і можуть бути навіть більш сприятливими в температурному відношенні. Влітку мікроклімат жилих приміщень здебільшого малосприятливий.
Отже, вибір житла для днювань залежить від багатьох моментів: пори року, кліматичних і мікрокліматичних умов. До цього слід додали розміщення хлівів відносно житла і відстань житла від місця виплоджування комара.
Не менш істотним фактором є місце перебування здобичі, тобто великої худоби, а також людини. При відсутності великих тварин, навіть так звані «дикі» види (An. hyrcanus, An. bifurcatus) можуть залітати в найближчі житла людини, так само, як Anopheles maculipennis переміщується за скупченням людей і худоби в поля і степові пасовища.
Потреба в кровоссанні у самиці анофелес залежить від двох моментів: від потреби живлення кров'ю під час дозрівання яєць, яким необхідно залізо крові, а по-друге, для кращого розвитку жирового тіла у період підготовки до зимівлі.
Епідеміологічне значення самиць комарів, як тимчасових паразитів, визначається ступенем контакту з людиною.
Частота нападів самиці анофелес залежить від температури повітря, поскільки це стимулює активність комара і визначає тривалість перетравлювання кров'яної їжі. Влітку в оптимальних температурних умовах процес перетравлювання крові супроводжується і закінчується дозріванням яєць, тому що самиця, що перетравила кров, відразу стін здатною відкладати-яйця (гонотрофічна гармонія). В цей час гопотрофічний цикл триває від 2 до 3 діб. Після підкладання яєць самиця знову нападає на здобич, тому що для розвитку наступної порції яєць потрібний новий прийом крові. Отже, влітку самиця може нападати на здобич кожні 4 — 5 діб.
В деяких місцевостях комарі нападають переважно на людину, а и інших — на велику худобу. Якщо має місце останнє явище, то при значній кількості переносників малярії тут фактично відсутня (анофелізм без малярії). Останнім часом (1920 рік) це явище було причиною появи ряду гіпотез; тут слід відзначити гіпотезу Рубо (Roubaml) про зоофільні і антропофільні раси, що нині має лише історичне значення.
Гіпотеза Рубо була поштовхом до вивчення генотипового складу популяції An. maculipennis, причому з'ясувалось, що цей вид є збірним і складається з багатьох підвидів (рас) (Фаллероні, Свелленгребель та його школа). Деякі з цих рас через свої біологічні особливості є переважними або єдиними переносниками, так що анофелізм без малярії ряд авторів пояснює відсутністю цих рас.
Однак, апофелізм без малярії являє собою, очевидно, складне явище, яке не може скрізь викликатися тими ж самими причинами, в тому числі і расовим складом малярійних комарів.
Щодо існування «зоофільних рас», які віддають перевагу крові великих тварин, то це можна пояснити легкістю виявлення здобичі, її доступністю і відносною чисельністю порівняно з людиною.
Для кожного окремого малярійного вогнища потрібне старанне вивчення всіх обставин, що роблять людину об'єктом переважного нападу анофелес. Дуже часто ці обставини можна усунути при організації відповідних заходів для відтягання комарів від людини (зоопрофілактика, раціональне планування населених пунктів, раціональне збудування жилих приміщень).
В епідеміології малярії істотне значення має тривалість життя літньої самиці, тому що збільшення його підвищує можливість нападу самиці на гаметоцитоносіїв і збільшує її небезпеку як переносника малярії.
У природних умовах літня самиця живе в середньому коло одного місяця, однак цей строк є достатнім не тільки для повного циклу спорогонії, а й для повторних нападів на людину.
В лабораторних умовах самиця комара може прожити значно довше, тому що загибель її в природних умовах слід вважати передчасною. Дуже небезпечні моменти для самиці: необхідність перельотів до водойм для відкладання яєць, самий процес відкладання і повторний напад на здобич.
В ці моменти самиця підпадає під вплив ряду несприятливих зовнішніх моментів, починаючи від метеорологічних умов і закінчуючи загибеллю від природних ворогів.
Разом з тим ряд екологічних факторів іноді є сприятливим для збільшення тривалості життя самиць анофелес у період їх активної життєдіяльності. В цьому випадку збільшення кількості гаметоцитоносіння призводить до спалахів малярії.
Нарешті, в епідеміологічному відношенні мають значення зміни поведінки комара в період підготовки його до зимівлі. При цьому спостерігаються при ссаннях крові, які тривають, або сповільнення у дозріванні яєць, або повне пригнічення функції яєчників (гонотрофічна дисоціація). Гонотрофічна дисоціація призводить до того, що комар залишається в приміщенні із здобиччю, тому що у нього зникає стимул вильоту для відкладання яєць. Це явище спостерігається наприкінці літа і на початку осені, коли до того ж, як описано вище, мікрокліматичні умови у житлах стають більш сприятливими.
Ця тривала затримка самиць комарів у жилих приміщеннях, куди починають збиратися малярійні комарі з природних сховищ, призводить до підвищення можливості зараження людини малярією, причому один заражений комар здійснює внутрішньородинне зараження.
Ці дані визначають і характер боротьби з комаром на днюваннях: влітку (період гонотрофічної гармонії) треба зосереджувати увагу на садибах і жилих приміщеннях маляриків, проводячи обробки через більш короткі проміжки часу, ніж тривалість перетравлювання самицею крові; Наприкінці літа и особливо на початку осені боротьбу треба провадити в усіх жилих приміщеннях із щоденними обробками, особливо в місцях масового скупчення комарів.
На закінчення слід зазначити, що для розвитку малярії, в якій-небудь місцевості обов'язкові такі умови:
Похожие работы
... лька десятків років тому. 4. Микола Федорович Гамалія Велика заслуга в розвитку імунології, теорії інфекції належить М.Ф. Гамалії, відкрившому також явище бактеріофагії. Микола Федорович Гамалія – видатний вчений, мікробіолог, епідеміолог, інфекціоніст. Народився 5 лютого 1859 року в Одесі. Закінчив Новоросійський університет в Одесі (1880 рік), а в 1883 році, по закінченню Воєнно-медичної ...



... У сухому, захищеному від світла місці. Синоніми: Білтрід, Цезол, Цестокс, Цистріцид, Дронцит, Піквітон, Азінокс, Балтріцид. Хлоксил (Chloxylum) Фармакологічна дія: Антигельмінтний засіб. Показання до застосування: Гельмінтози печінки (хвороби печінки, що викликаються паразитарними червами: опісторхоз, фасціольоз, клонорхоз). Спосіб застосування: Цикл лікування хлоксилом продовжується ...
... і співдоповідей, розподілити завдання, повідомити питання, що будуть обговорюватися на семінарі. Крім тих методів опитування, що практикуються на основних предметах біологічного циклу, на факультативних заняттях можна застосовувати залікові заняття, що дозволяють забезпечити фронтальну перевірку всіх учнів факультативної групи. Ці заняття звичайно проводяться після вивчення чергової теми. Вони ...
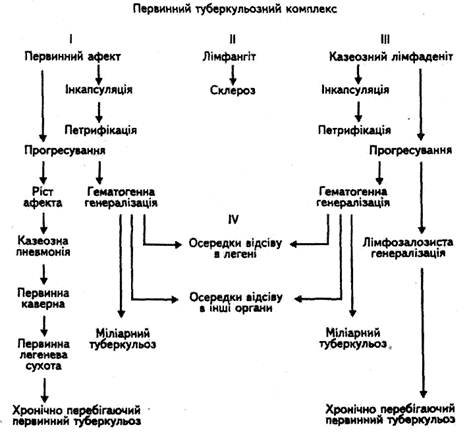
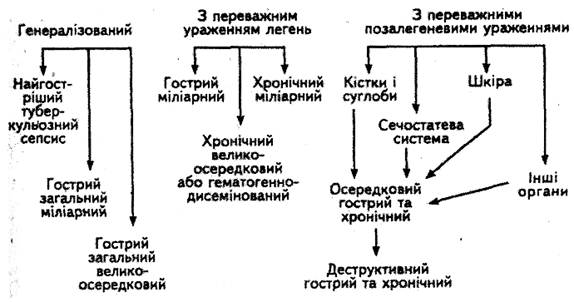
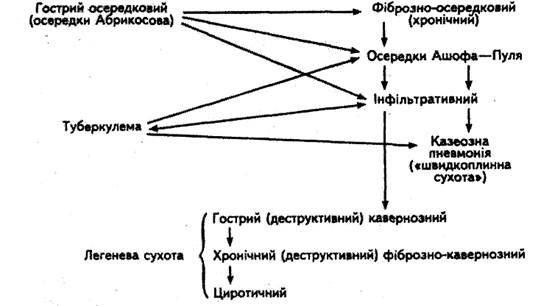

... межах господарства, заготівлею та реалізацією тваринницької продукції. Ферми і приватні господарства комплектують лише здоровими, перевіреними на туберкульоз тваринами, з благополучних щодо інфекційних захворювань господарств. У благополучних щодо туберкульозу господарствах потрібно забезпечувати нормативні умови утримання, годівлі та використання сільськогосподарських тварин, а також ізольоване ...
0 комментариев