Навигация
4. Зберігання інформації
Зберігання інформації пов'язане з переходом короткочасної і проміжної пам'яті в довготривалу пам'ять. Процес переходу інформації з короткочасної в довготривалу пам'ять називається консолідацією пам'яті. При цьому пам'ять набуває стійкої форми. Вона не змінюється в часі а також при додаванні нової інформації.
5. Довготривала пам'ять
Довготривала пам'ять визначає збереження раніше отриманої інформації протягом довгого часу. Процеси фіксації слідів в довготривалій пам'яті здійснюються краще при повторних діях. Найшвидше процес консолідації пам'яті відбувається при дії емоційно значущих подразників. Довготривала пам'ять по своєму механізму якісно відрізняється від короткочасної пам'яті і не порушується при таких екстремальних діях на мозок, як механічна травма, електрошок, наркоз і ін.
Механізм довготривалої пам'яті остаточно не встановлений. Декілька теорій з різних позицій пояснюють механізми довготривалої пам'яті.
Морфологічні теорії:автори цих теорій вважають, що довготривала пам'ять пов'язана з утворенням нових синаптичних контактів на тілах нейронів ЦНС, а також зі збільшенням розмірів синапсів. Вважають, що довготривала пам'ять також пов'язана з розростанням дендритів і збільшенням числа шипів на дендрітному дереві нейронів мозку. Передбачають, що при цьому збільшується число колатералів аксонів нейронів.
Довготривалу пам'ять також пов'язують із зміною молекулярних структур нейронів – збільшенням числа мікротрубочок і інших молекулярних утворень. У всіх цих випадках збільшується число нових терміналів на нейронах мозку, що сприяє поширенню по структурах мозку більшої інформації.
На відміну від концепцій довготривалої пам'яті, пов'язаних з формуванням нових синаптичних контактів між нейронами, канадський вчений Хебб висунув гіпотезу, згідно якої довготривала фіксація слідів пам'яті пов'язана із стійкими змінами синаптичної провідності в межах існуючих пулів синапсів. Ця точка зору лягла в основу багатьох сучасних уявлень про молекулярні механізми пам'яті.
Каскадні зміни в синапсі при формуванні пам'яті. Навчання запускає каскад біохімічних процесів в мозку. Відбуваються зміни використання глюкози, посилюються взаємодії між нейромедіаторамі і їх рецепторами, що веде до зміни властивостей синаптичних мембран і підвищення ефективності зв'язку між пре- і постсинаптичними нейронами. Ці зміни у свою чергу породжують сигнали для клітинного ядра, в якому спочатку активується декілька "раніх" генів, а потім і гени необхідні для синтезу нових компонентів синаптичних мембран, особливо глікопротєїнів, які в подальші години включаються в синаптичні мембрани, збільшуючи зони синаптичного контакту і число шипів на дендрітах.
Гліальна теорія: Автори цієї теорії вважають, що довготривала пам'ять пов'язана з активністю гліальних клітин, що оточують нейрони. Ці клітини у міру навчання тварин синтезують спеціальні речовини, що полегшують синаптичну передачу, а також змінюють збудливість відповідних нейронів. При навчанні в гліальних клітинах збільшується вміст РНК.
Встановлено, що деполяризація нейронів викликає їх мієлінізацію, що також приводить до зростання ефективності синаптичної передачі збуджень. Деякі автори вважають, що клітини глії своєрідно програмують діяльність нейронів мозку.
Медіаторна теорія: Показано, що під впливом навчання в синапсах ЦНС збільшується кількість холінорецепторов. При цьому підвищується чутливість нейронів мозку до мікроіонофоретичному підведенню ацетілхоліну. Антагоністи ацетілхоліну, навпаки, порушують навчання і відтворення, викликають амнезію. Встановлено також, що навчання тварин на основі електрошкірного підкріплення супроводжується активацією норадренергічних механізмів, а навчання на основі харчового підкріплення знижує рівень норадреналіну в мозку тварин. Зниження рівня норадреналіну в мозку за допомогою фармакологічних речовин також уповільнює навчання і викликає амнезію. При цьому достовірно порушуються процеси відтворення слідів пам'яті. Показана участь дофаміна в механізмах пам'яті. У процесах, пов'язаних з консолідацією пам'яті, беруть участь серотонінергічні механізми. Серотонін бере участь в процесах навчання на емоційно позитивному підкріпленні, порушуючи виконання у тварин оборонних навиків.
Вважають, що активація холінергічних синапсів викликає конформаційні перебудови постсинаптичних мембран, що підвищують синаптичну провідність.Моноамінергічні механізми, пов'язані з підкріпленням, активують внутрішньоклітинні постсинаптичні процеси за участю циклічних нуклеотидів – цАМФ і цГМФ. В результаті подальших метаболічних внутрішньоклітиних процесів синтезуються спеціальні білкові молекули, які у свою чергу стабілізують первинні зміни синаптичних мембран. В результаті в структурах мозку формуються зони підвищеної синаптичної провідності, що і визначає формування відповідних енграм пам'яті.
Підтвердженням такої точки зору є концепція, запропонована Г.Лінчем і М.Бодрі, яка виходить з того, що повторна стимуляція нейрона приводить до збільшення вмісту іонів кальцію в постсинаптичній мембрані. Це у свою чергу активує фермент – кальцийзалежну протєїназу, яка розщеплює один з білків мембрани, тим самим приводячи до звільнення раніше неактівних білків глутаматних рецепторів. Їх число зростає, внаслідок чого збільшується провідність синапсів.
У механізмах синаптичної пам'яті беруть участь також гама-аміномасляна кислота, глутамінова кислота, проте механізми їх дії у багатьох відношеннях все ще залишаються невивченними.
Молекулярні теорії:більшість молекулярних теорій пов'язує механізм довготривалої пам'яті з діяльністю генетичного апарату нейронів і гліальних елементів мозку, зокрема з синтезом РНК.
Роль РНК. Встановлено, що нейрони мозку в порівнянні з клітинами інших тканин організму містять максимальну кількість РНК – 20-2000 пг на кожну нервову клітину, що складає 5-10% її сухої маси. Крім того, нейрони характеризуються максимальною кількістю активних генів.
Для з'ясування ролі РНК в механізмах пам'яті щурам вводили 8-азагуанін, блокуючий приєднання гуаніна і РНК, що тим самим спотворює синтез. 8-азагуанін погіршував формування умовних рефлексів. В той же час його введення після вироблення умовних рефлексів не впливало на їх прояв. Звідси витікає, що РНК має переважне значення для вироблення навиків, а не для їх зберігання.
Введення щурам блокатора синтезу ДНК-азітотімідина перешкоджає переходу короткочасної пам'яті в довготривалу.
Хіден передбачив, що під впливом прихідної до нейронів імпульсациі відбувається перегруповування основ в молекулах РНК що у свою чергу приводить до синтезу на таких змінених молекулах РНК специфічного білка, що обумовлює вибірну чутливість нейронів саме до даної конфігурації імпульсів.
Підтвердженням такої точки зору є експерименти, що показали, що екстракти, позбавлені РНК, мали аналогічну дію. Звідси виникло припущення про перенесення пам'яті "специфічними білками".
Білки пам'яті.Про участь білкового синтезу в консолідації енграм довготривалої пам'яті свідчать експерименти з введенням блокаторів синтезу білка.
Введення навченним тваринам пуроміцина, що викликає передчасне зняття білка з рібосом, зменшує час зберігання вироблених навиків. Введення пуроміцина до навчання не впливає на швидкість вироблення навиків, тобто на короткочасну пам'ять.
Блокада синтезу білка на стадії трансляції на рібосомах циклогексимідом і іншими антибіотиками або на стадії транскрипції – актіноміцином D також пригнічує раніше вироблені навики у тварин. При пригніченні білкового синтезу при короткочасному навчанні спостерігається збереження вироблених навиків лише протягом декількох хвилин або години після навчання. Через 1 – 2 години після введення блокаторов синтезу білка спостерігаються глибокі порушення в збереженні вироблених навиків. Все це свідчить про те, що процеси білкового синтезу неефективні найближчим часом після навчання. Вони включаються пізніше і ведуть до формування енграми довготривалої пам'яті.
Деякі з білків пам'яті – білок S-100, 14-3-2; холінорецептивний білок ацетілхолінестераза - вже виділені з мозку. Білок S-100 активно взаємодіє із зовнішньою мембраною і скоротливими мембранами нейрона за участю іонів кальцію. Білок 14-3-2 є ензимом, що бере участь в реакціях гліколізу у нейронах. Встановлено, що в процесі навчання білок S-100 переважно накопичується в гипокампці, а білок 14-3-2 – у корі мозку. Деякі автори розглядують білок S-100 як гліальний білок.
Нейропептиди. Деякі фрагменти АКТГ беруть участь в механізмах пам'яті. Видалення у тварин гіпофіза істотно порушує у них прояв раніше вироблених навиків. Тваринні з природженим дефектом вироблення вазопрессина не здатні до утворення захисних навиків. Вироблення навику відновлюється у цих тварин лише при додатковому введенні вазопрессина. Характерно, що при цьому у тварин страждає не сам процес навчання, а саме консолідація енграм, що сформувалися.
Інший олігопептид – окситоцин – порушує у тварин збереження вироблених навиків незалежно від типа навчання.
На навчання і пам'ять впливають інші ендогенні олігопептиди, наприклад ендорфіни і енкефаліни. Вони уповільнюють затухання умовних рефлексів, покращують їх збереження, хоча і погіршують їх формування.
Нейропептиди в механізмах пам'яті тісно взаємодіють в синапсах з медіаторами; після навчання кругообіг катехоламінів в мозку збільшується.
Похожие работы
... : - приступоподібна шизофренія, шизоафективний варіант, депресивний тип; - шизоафективний психоз, депресивний тип; - шизофреноформний психоз, депресивний тип. РОЗДІЛ 2. ЗМІНИ ОСОБИСТОСТІ ПРИ ШИЗОФРЕНІЇ 2.1 Дитяча та юнацька шизофренія Шизофренічний процес у дитячому і підлітковому віці тісно зв'язаний із критичними періодами, кожний з яких відрізняється своєю специфікою, ...
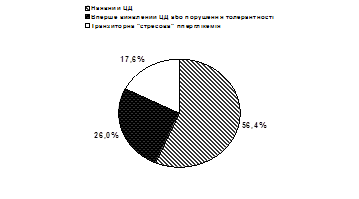
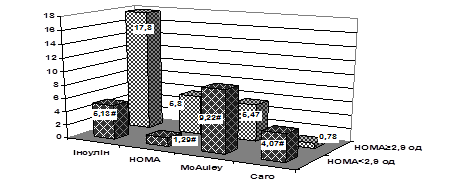
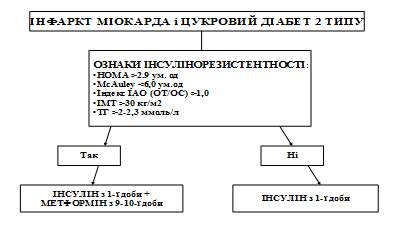
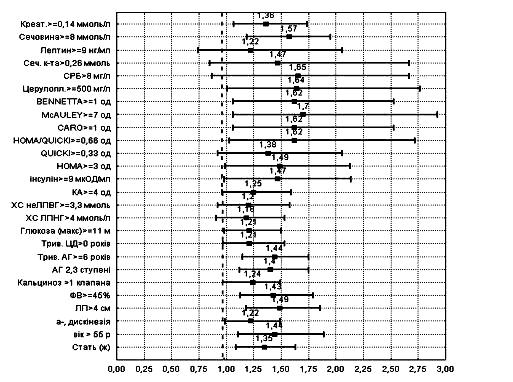
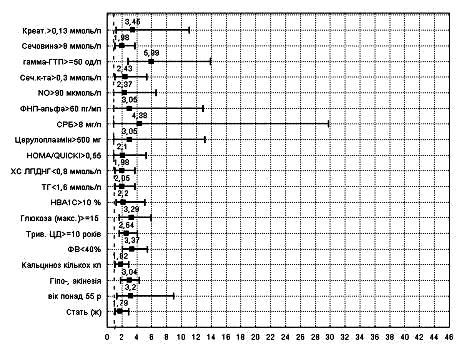
... ія та ендокринна хірургія. – 2004. – № 1. – С. 45-51 (Здобувач провів клінічні дослідження, узагальнення отриманих результатів, сформулював висновки). 7. Скибчик В.А., Соломенчук Т.М. Особливості перебігу інфаркту міокарда у хворих на цукровий діабет ІІ типу залежно від статі та віку // Український медичний часопис. – 2004. – № 2. – С. 115-118 (Здобувач провів клінічні дослідження, узагальнення ...
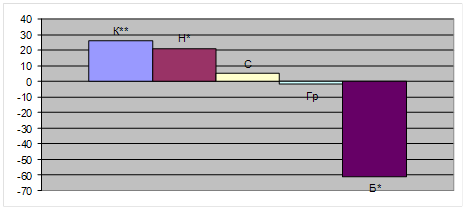
... основних характеристик збудливості сегментарних моносинаптичних рефлекторних систем у білих щурів; провела огляд літератури). АНОТАЦІЯ Гузь Л.В. Нейрометаболічні особливості та інтегративна діяльність центральної нервової системи за умов експериментальної гіпертироксинемії у щурів. – Рукопис. Дисертація на здобуття наукового ступеня кандидата біологічних наук за спеціальністю 03.00.13 – ...
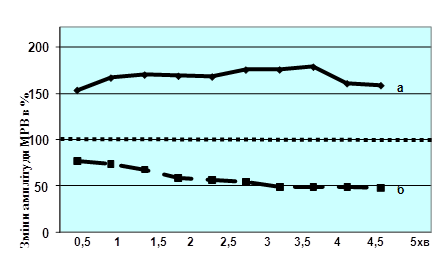
... Т.Г. – 3.№ 200605148; Заявл. 10.05.06; Опубл. 15.11.06, бюл. 11. (Дисертантом взято участь у зборі матеріалів та обробці отриманих даних). АНОТАЦІЯ Перцева Т.Г. Особливості перебігу дисциркуляторної енцефалопатії у хворих з цукровим діабетом II типу. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.01.15 – нервові хвороби. – ДУ “Інститут ...
0 комментариев