Навигация
Белок переносчик вместе со связанным лигандом перемещается с одной стороны мембраны на другую в результате физической диффузии
27693
знака
0
таблиц
0
изображений
1. Белок переносчик вместе со связанным лигандом перемещается с одной стороны мембраны на другую в результате физической диффузии.
2. Молекулы переносчики вероятно белки фиксированы на мембране, но могут претерпевать конформационные изменения чтобы обеспечить перенос субстрата через мембрану.
Комплексные соединения: один из ионов, обычно положительно заряженный – в центре - центральный ион. Вокруг расположено некоторое число противоположно заряженных ионов или электронейтральных молекул, называемых лигандами.
Аквакомплексы – лиганд вода.
Гемоглобин – гем- центральный ион – ион железа – вокруг него координированы четыре атома азота, принадлежащих к сложному лиганду с циклическими группировками.
Согласно одному из предположений, связывающий белок обладает сильным сродством к транспортируемому веществу (субстрату) и прочно связывается с ним на наружной поверхности клетки. Образовавшийся комплекс белок-субстрат далее диффундирует к внутренней стороне мембраны. Здесь в результате процесса, сопряженного с самопроизвольно протекающей экзергонической реакцией, например, с гидролизом АТФ, конформация белка меняется таким образом, что его сродство к субстрату уменьшается. В результате транспортируемое вещество переходит в клетку, а связывающий белок диффундирует обратно к наружной поверхности. Там его конформация возвращается к исходной, вероятно, под влиянием химических воздействий.
Какие классы молекул и ионов транспортируются через мембраны? Из окружающей среды в клетку поступают неорганические ионы, причем в результате переноса иногда они сильно концентрируются. Клетки слизистой оболочки желудка могут концентрировать ионы водорода в желудочном соке примерно до 0,16 М. В клетки животных и человека активно транспортируются аминокислоты. Обычно наряду с системой, для которой характерны высокое сродство к аминокислоте и способность “перекачивать” из области с очень низкой концентрацией, существуют параллельно функционирующие системы с рецепторами не обладающими столь высоким сродством к субстрату. Сахара в этих системах проходят через внутреннюю мембрану в виде фосфатных эфиров (групповая транслокация).
В другой системе транспорт аминокислот и лактозы сопряжен с системой переноса электронов связанной с мембраной окислительно-восстановительной цепи. Считают, что эта система независима от синтеза АТФ.
Внутриклеточные органеллы имеют собственные системы, концентрирующие ионы. Так митохондрии могут концентрировать ионы калия, кальция, магния и других двухвалентных металлов, а также и дикарбоновые кислоты. У митохондрий транспорт многих веществ происходит скорее всего за счет обменной диффузии, т.е. путем вторичного активного транспорта.
Генетические аспекты транспорта.
У человека описан целый ряд заболеваний, связанных с дефектами мембранного транспорта. При некоторых таких заболеваниях нарушаются реабсорбция веществ в почечных канальцах и процесс всасывания в тонком кишечнике. Цистонурия – наблюдается образование камней из цистина в почках и мочевом пузыре. Известны также случаи выделения больших количеств аргинина, лизина и орнитина.
Существование подобных наследственных заболеваний свидетельствует о том, что и у человека клетки обладают способностью концентрировать различные АК и другие вещества.
В клетках почечных канальцев вещества поглощаются на одной стороне клетки и выделяются в кровоток с другой ее стороны.
Еще одно редко встречающееся нарушение абсорбционных процессов у человека приводит к развитию почечной гликозурии. В этот процесс вовлечены проксимальные почечные канальцы.
Натрий, калий и кальциевый насосы.
Почти во всех клетках концентрация натрия сравнительно невелика, тогда как калия – достаточно высока.
Согласно одной из гипотез, цитоплазма по своим свойствам напоминает ионообменные смолы, несущие фиксированные заряды. Ионообменные смолы с большим числом сшивок избирательно связывают определенные ионы. Например, смолы, содержащие остатки сульфокислот, связывают калий, а смолы, содержащие остаток фосфорной кислоты – натрий.
В настоящее время наиболее общепринятой является не ионообменная гипотеза, а гипотеза существования в клетках ионного насоса, выкачивающего из клеток ионы натрия и накачивающего в них ионы калия.
Оказалось, что перенос ионов блокируется ингибиторами, например, цианидом, который, как известно, нарушает почти все процессы окислительного метаболизма в клетках. Однако блокирование цианидом снимается при добавлении к клеткам АТФ или других фосфатных соединений, характеризующихся высоким значением потенциала переноса групп. Поскольку в клетках эукариот самым активным соединением такого рода является АТФ, предполагается, что именно он служит естественным источником энергии при концентрировании ионов.
Поглощение клетками ионов калия и выделение ионов натрия специфически блокируется некоторыми сердечными гликозидами. Наиболее часто в этих целях используют уабаин. Меченый тритием уабаин связывается с наружной поверхностью клеток.
Установлено, что при избытке ионов натрия внутри клетки, а ионов калия во внешнем окружении происходит гидролиз АТФ. Скорость гидролиза определяется концентрацией ионов обоих щелочных металлов и числом мест, связывающих уабаин. Для этого процесса необходимо наличие ионов магния.
Рассматриваемые выше факты привели к концепции, предполагающей, что (натрий и калий) – зависимая АТФ-аза и является, по существу, мембранным ионным насосом. Для активации ферментной системы ионы калия и натрия должны находиться по разные стороны от мембраны.
Стехиометрические соотношения в натрий-калиевом насосе весьма своеобразны. При распаде каждой молекулы АТФ из клетки выкачивается 3 иона натрия, а извне в клетку накачиваются 2 иона калия. Поскольку из клетки выкачивается больше положительно заряженных ионов , чем попадает в нее, внутри клетки создается избыточный отрицательный заряд. Наличие отрицательного заряда внутри клетки было установлено уже давно, путем измерения электрического мембранного потенциала. Поскольку клеточная мембрана все же проницаема для ионов калия, возникновение мембранного потенциала приводит к диффузии этих ионов через мембрану внутрь клетки, что обусловливает частичную нейтрализацию отрицательного заряда на мембране. Когда скорость пассивной диффузии уравновешивает мембранный потенциал, образующийся за счет активного транспорта устанавливается стационарное состояние.
Таким образом, достоверно установлено, что транспорт натрия и калия ионным насосом обеспечивается энергией гидролиза АТФ.
Процесс расщепления АТФ должен быть сопряжен с каким-то конкретным механизмом, обеспечивающим перекачку ионов, который до сих пор остается невыясненным.
Предполагается, что белки ионного насоса находятся в двух конформациях. В одной конформации (А) белок прочно связывает 3 иона натрия, а в другой (В) – 2 иона калия. АТФ играет роль “мотора”, осуществляющего конформационные изменения белка и кроме того, может принимать непосредственное участие в формировании участков связывания ионов.
Самопроизвольное связывание ионов натрия запускает реакцию фосфорилирования, в результате которой фосфорильная группа комплекса магний – АТФ – переносится и фосфорилирование в свою очередь инициирует переход белка в конформационное состояние В, при котором канал становится открытым с наружной стороны и закрытым – с другой. Одновременно уменьшается сродство по отношению к ионам натрия и эти ионы выходят наружу. На следующем этапе связываются 2 иона калия. Для конформации В характерно высокое сродство к ионам калия. Обратный переход в конформацию А, сопровождающийся освобождением ионов калия во внутреннее пространство клетки, инициируется гидролитическим отщеплением фосфорильной группы в виде неорганического фосфата. Химическая природа участков связывания ионов натрия и калия в ионном насосе неизвестна.
Проведение нервных импульсов.
Способность мембран поддерживать разность концентраций ионов между цитоплазмой и внеклеточной средой обуславливает их возбудимость, что имеет важное практическое значение. Разделение зарядов создает как бы “аккумулятор” легко доступной энергии, которая может быть использована для проведения электрических сигналов через клеточную мембрану. В настоящее время мало известно о химической природе проведения нервных импульсов, однако электрические характеристики этого процесса детально изучены и описаны.
Если ввести в клетку через мембрану микроэлектрод, то можно измерить разность потенциалов между внешней средой и содержимым клетки. Эта разность потенциалов между внешней средой и содержимым клетки получила название потенциала покоя. Своим происхождением она обязана различиям в концентрации ионов. Если на каком-либо участке мембраны проницаемость для ионов натрия увеличивается, то эти ионы устремляются внутрь клетки, нейтрализуя ее отрицательный заряд. Клеточная мембрана при этом деполяризуется.
В большинстве аксонов нервных клеток используется более эффективный способ проведения импульса, основанный на развитии потенциала действия. Потенциал действия _ это импульс, проходящий вдоль аксона и специфически изменяющий за доли секунды мембранный потенциал. Исходный отрицательный потенциал быстро падает до нуля, затем достигает положительного значения, после чего снова устанавливается потенциал покоя.
Отличительным свойством нервных волокон, обладающих способностью проводить импульсы, является наличие миелиновой оболочки.
Миелиновые мембраны надежно изолируют аксон от окружающей среды на всем протяжении, кроме специальных участков, перехватов Ранвье. За счет этого нервный импульс обретает способность как бы перескакивать с одного участка на другой.
Поскольку число каналов, проводящих ионы мало, скорость прохождения натрия через открытые каналы должна быть высокой. Отсюда следует, что эти каналы не могут работать с помощью переносчиков, а должны представлять собой просто “поры”, открывающиеся и закрывающиеся в соответствии с изменениями мембранного потенциала.
Подобно ионам натрия и ионы кальция активно выводятся из клетки. Градиент концентрации ионов кальция по разные стороны от мембраны поддерживается при помощи кальциевого насоса. Работе насоса противодействует очень медленная диффузия ионов внутрь клетки.
Транспорт через митохондриальные мембраны.
Митохондриальные мембраны отличаются высокой избирательностью. Некоторые неионизированные вещества легко проходят через нее, тогда как транспорт ионных веществ, включая анионы дикарбоновых и трикарбоновых кислот находятся под жестким контролем. В некоторых случаях анионы перемещаются в результате энергозависимого “активного транспорта”. В других случаях анион может пройти внутрь лишь в обмен на другой анион, выходящий наружу. Во всех этих случаях необходимо участие специфических транслоцирующих белков-переносчиков.
Одна из систем транслокации производит обмен АДФ на АТФ.
Отдельный переносчик ведает доставкой фосфора. Пируват тоже попадает в митохондрии с помощью собственного переносчика.
С другой стороны, анион дикарбоновых кислот, малат и альфа-кетоглутарат обмениваются в соотношении 1:1, равно как и аспартат и глутамат.
Мембраны митохондрий непроницаемы для НАДН. У животных восстановленные эквиваленты НАДН поступают в митохондрии косвенным путем. Эту функцию выполняет сложный малат-аспартатный челночный механизм.
Существует предположение, что вывод аспартата из митохондрии связан с потреблением энергии, и в этом случае можно провести аналогию с работой натриевого насоса в цитоплазматической мембране
Похожие работы
... – высокоаэробных.[6,2000] Показано, что ряд дегидрогеназ используют только НАД или НАДФ, другие могут катализировать окислительно-восстановительные реакции в присутствии любого из них. В процессе биологического окисления НАД и НАДФ выполняют роль промежуточных переносчиков электронов и протонов между окисляемым субстратом и флавиновыми ферментами. Основными источниками никотиновой кислоты и ее ...
... формируются вдоль плазматической мембраны при захвате ЛП клетками с помощью специфических апо-В,Е-рецепторов. Этот путь транспорта ЛП направлен на обеспечение внутренних потребностей клеток в холестерине и не приводит к поступлению липопротеидных частиц в интиму артерий. Окаймлённые эндоцитозные везикулы, содержащие липопротеидные частицы, вступают в контакт и сплавляются с лизосомами, где ЛП ...
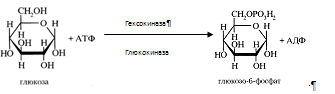
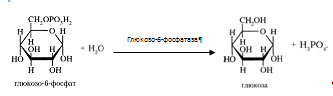
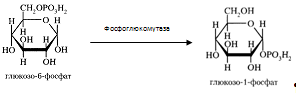

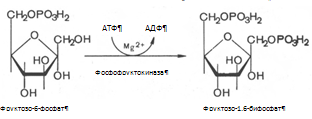
... 3) гепатодепрессии (печёночно-клеточной недостаточности, малой недостаточности печени, недостаточности синтетических процессов); 4) воспаления; 5) шунтирования печени; 6) регенерации и опухолевого роста. При подозрении на конкретную патологию учитываются основные биохимические синдромы, характерные для данного заболевания. За основу берётся стандартная программа функционального обследования, но ...
... кислоты, которые относят к незаменимым жирным кислотам (линолевая, линоленовая, арахидоновая), которые не синтезируются у человека и животных. С жирами в организм поступает комплекс биологически активных веществ: фосфолипиды, стерины. Триацилглицеролы – основная их функция – запасание липидов. Они находятся в цитозоле в виде мелкодисперсных эмульгированных маслянистых капелек. Сложные жиры : ...
0 комментариев