Навигация
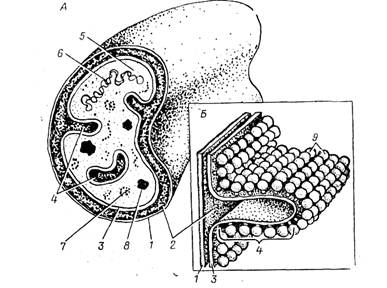
Характеристика метаболізму та життєвий цикл хламідій
50224
знака
3
таблицы
7
изображений
4. Характеристика метаболізму та життєвий цикл хламідій
4.1 Характеристика метаболізму хламідій
Як вже згадувалось, хламідії, як енергетичні паразити живих епітеліальних клітин, використовують для свого метаболізму їх АТФ. В даний час аналіз генома показав, що хламідії здатні синтезувати АТФ, хоч і в незначних кількостях, шляхом гліколізу і розщеплювання глікогену [4].
Як було виявлено Weiss і співавт. (1964), у хламідій спостерігається анаеробний метаболізм глюкози, що проходить по шляху пептозофосфату і одного з гліколітичних шляхів. Тобто, при додаванні кофакторів яких вони потребують, хламідії можуть метаболізавати глюкозу, а також піровиноградну і глутамінову кислоти [7].
Ця активність здійснюється на низькому рівні і за цих умов приводить до втрати АТФ і НАД. Система транспорту електронів відсутній, хоча обидві частинки містять цитохром С-редуктазу. Макромолекулярний синтез клітин хазяїна гальмується хламідіями, і високоенергетичні субстанції кліток хазяїна переходять на синтез протеїнів і ліпідів хламідій. Таким чином, забезпечення мікроорганізму метаболітами здійснюється в основному за рахунок життєдіяльності клітин хазяїна. Деякі з цих метаболітів (ізолейцин) можуть бути інгібіторами зростання хламідій і, ймовірно, можуть мати відношення до латентної течії при хламідіозі [Hatch, 1975] [2].
Як яскравий приклад метаболічної активності хламідій і впливу на метаболізм клітин хазяїна є дослідження Харківського ПІІ дерматології і венерології на споживання глюкози клітинами [17]. Досліди проводили на зараженій культурі клітин L-929. Утилізація глюкози інфікованою культурою достовірно вище протягом всього процесу культивування. Оскільки в інфікованій культурі інтенсивне зростання кліток не відбувається, можна припустити, що збільшення споживання глюкози кліткою пов'язано з цитоплазматичним включенням, що розвивається, тобто хламідійною інфекцією [18]. Через 5 г після зараження, коли фагосома з елементарних тілець (ET) переміщається в пластинчастий комплекс і ЕТ перетворюється на ретикулярні тільця (РT) через проміжні тільця (PT), споживання глюкози клітками культури більш ніж в 7 разів вище, норми. Можливо, в цей період активізуються біосинтетичні процеси клітини-хазяїна, продукти яких будуть використані збудником на побудову включення.
Згідно з літературними даними [19], починаючи з 10-12 г після впровадження збудника і до 36 г РT, зазнає 10-11 поділень, при цьому утворюється пул кліток, що є цитоплазматичним включенням. Звичайно цей процес закінчується до 24 г. Можливо, достовірне зниження споживання глюкози інфікованою клітиною, яке більш ніж в 4 рази вище в порівнянні з нормою, в цей період пов'язано з уповільненням її метаболічній активності при швидкому зростанні морфологічних структур збудника.
Можна припустити, що перетворення метаболічно активних РT в неактивні ЕТ, як правило, що завершується до 48 г, супроводжується розкріпаченням власних метаболічних процесів, клітини, і, як наслідок, спостерігається тенденція до збільшення споживання глюкози [18].
До 72 г цикл розвитку хламідій завершується. При мікроскопуванні препаратів спостерігали зрілі бочкоподібні включення збудника, що займають практично весь об'єм клітини. Значна частина моношару зруйнована, вміст глюкози в середовищі збільшений. І не дивлячись на це, споживання глюкози інфікованою клітиною в цей період в 3 рази вищий, норми.
Таким чином встановлено, що споживання глюкози інфікованою кліткою впродовж всього циклу розвитку збудника достовірно вищий, ніж у нормі. Спостерігаються певні кореляційні залежності між стадіями розвитку збудника і споживанням глюкози інфікованою кліткою.
Також проводилися дослідження синтезу білка в клітинах хазяїна по активності трансаміназ. Активність аланіномінотрансферази (AлT), аспартатаминотранферази (AсT) визначали по методу С. Райтмана, С. Френкеля (1957).
Так, до 24 г активність обох трансфераз у сфері культивування нормальних клітин достовірно збільшилася (AсT - в 4 рази, АлТ-в 8 разів) і залишилася на досягнутому рівні весь період, що залишився. Можна припустити, що інтенсифікація діяльності ферментів до 24 г культивування пов'язана з посиленим утворенням амінокислот в ході реакцій трансамінування, які використовуються як субстрат при синтезі білкових молекул в культурі, що активно ділиться (експоненціальна фаза зростання). До 72 г розвиток клітин в культурі виходить на плато. Крім того, спостерігається деяка тенденція до зниження активності ферментів [17].
Через 24 г після інфікування не було відмічено достовірних відмінностей між активністю ферментів в середовищі культивування нормальних клітин і в середовищі культивування інфікованих клітин, тоді як у всіх інших часових інтервалах активність ферментів в середовищі культивування інфікованих клітин була достовірно вища. Відносно низька активність ферментів в середовищі інкубації інфікованої культури клітин через 24 г може свідчити, про те, що амінокислоти інтенсивно включаються в синтез білка, і немає сенсу здійснювати ширше їх використання для інших цілей [18]. Разом з тим в період від 24 до 48 г культивування активність AлT і AсT збільшувалася майже в 2 рази, причому в цей період вона вища, ніж в нормі, відповідно в 2,9 і 3,5 рази. Це може бути пов'язане з інтенсифікацією процесу глюконеогенезу в інфікованій культурі. Дійсно, згідно з літературними даними [19]. саме у цей період завершується ділення РT, починається їх перетворення в ЕТ, а при мікроскопуванні в інфікованих клітках виявили гранули глікогену. Іншою причиною такого швидкого зростання активності ферментів в середовищі може бути збільшення проникності мембран інфікованої клітки.
До 72 г активність AлT зростає ще в 2,1 рази, в цей період вона вища, ніж в нормі, в 6,7 рази, AсT, відповідно, - в 5,6 рази. У цей період закінчується цикл розвитку хламідій, клітини руйнуються і росте вміст ферменту і середовища [17].
Таким чином, виявлені достовірні відмінності у ферментативній активності клітин нормальної і інфікованої культур. Показано, що активність ферментів в середовищі культивування інфікованих кліток збільшується в процесі дослідження, що корелює з циклом розвитку збудника [17].
Деякі факти, виявлені в ході дослідження, не знайшли свого пояснення у зв'язку з тим, що хламідії в процесі пристосування до внутріклітинного паразитизму виробили унікальні структури і біосинтетичні механізми, що не мають аналогів у інших бактерій. Не пояснений той факт, що у хламідій не виявлений висококонсервативний ген Ftsz, абсолютно необхідний для клітинного ділення всіх прокаріот, оскільки він відповідальний за утворення клітинної перегородки під час ділення клітки. У хламідії відсутній пептидоглікан - компонент клітинної стінки існує як у грампозитивних, так і у грамнегативних бактерій, але при цьому в геномі містяться гени, що кодують білки, які необхідні для його повного синтезу. Передбачається, що пептидоглікан, що синтезуються, або пептидогліконовий компонент мають інші функції, відмінні від інших бактерій [4].
Похожие работы

... а присутня тільки особлива система мембран, яка містить дві первинні дегідрогенази дихального ланцюга. Під дією кисню ці мембрани зливаються, утворюють складки і перетворюються в істинні мітохондрії, які містять цитохроми. РОЗДІЛ 2. МЕТОДИ ДОСЛІДЖЕННЯ МІТОХОНДРІЙ Значних успіхів було досягнуто у вивченні мітохондрій, при цьому значну роль зіграла електронна мікроскопія, яка дала можливість ...
... зі змістом білків 8 - 58 %, вуглеводів – 6 - 37 н жирів – 4 - 85 %. У залежності від умов вирощування в значній мірі змінюється також зміст вільних амінокислот, пігментів, вітамінів, мікроелементів. Водорості, особливо мікроскопічні, характеризуються найбільш високим КПД засвоєння світлової енергії в порівнянні з іншими фотосинтезуючими організмами. Багато видів здатні до ефективної утилізації сві ...
... синтезовані хімічним шляхом з'єднання, що не зустрічаються в природі, також атакуються мікробами. Всі викладені вище відомості, що становлять лише фрагменти того, що відомо сучасній мікробіології, та екології бактерії дають підставу зробити висновок про величезну й виняткову роль мікроорганізмів у круговороті речовин у природі. Види зелених водоростей та їх екологічне значення Зелені водорості ...
... Кролівництво та хутрове звірівництво кролі лисиця норка нутрія песець Рикетсійній кератокон'юктивіт Загальна інформація Рикетсійний кератокон’юнктивіт (keratokonynctivitis richetsion) – гостре інфекційне захворювання тварин з ураженням очей з розвитком катарального кон’юнктивіту і кератиту. Патогенез Не вивчено. Симптоми Інкубаційний період триває 10-12 діб. Перебіг хвороби гострий ...
0 комментариев