Навигация
Життєвий цикл хламідій всередині зараженої клітини
50224
знака
3
таблицы
7
изображений
4.2 Життєвий цикл хламідій всередині зараженої клітини
Зрілою формою збудника хламідіозу є спороподібне ЕТ з діаметром 250-300 нм, що володіє інфекційністю і обмеженою метаболічною активністю.
Ретикулярні тільця утворюються в процесі розмноження мікроорганізму всередині клітини-хазяїна, є попередником нового покоління ЕТ і є вегетативною формою хламідії. У протилежність ЕТ - вельми лабільні, неінфекційні і володіють вираженою метаболічною активністю. Проміжні тільця утворюються на двох стадіях циклу розвитку хламідій: на ранній, при перетворенні ЕТ в РT, і на пізніше, при реорганізації РT в ЕТ .
Цикл розмноження хламідій - облігатних внутріклітинних паразитів - реалізується при їх взаємодії з чутливою клітиною-хазяїном.
Схематично і спрощено основні морфологічні етапи цього циклу представлені на (рис.4.2.1). Інфекційні ЕТ адсорбуються на поверхні клітини, яка потім поглинає їх шляхом ендоцитозу. Доля фагоцитованих ЕТ на цьому першому критичному етапі взаємодії з клітиною-хазяїном може скластися двояко:
• EТ гине під впливом лізосомної активності клітини-хазяїна
• ЕТ зберігає життєздатність і вступає в цикл розвитку [5].
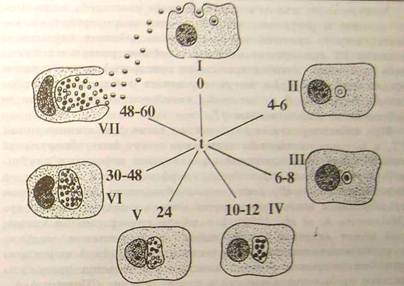
Рисунок 4.2.1 - Схема циклу розвитку хламідій в чутливій клітині: (Шаткін А.А., Мавров І.І., 1983) [5]
Хламідії здатні якимсь чином стимулювати специфічний фагоцитоз, що дає очевидні переваги облігатному внутріклітинному паразиту. Проникнення елементарних тілець в клітину супроводжується руйнуванням оболонок, внаслідок чого відбувається розм'якшення клітинних стінок. Присутність інфекційних частинок у фагосомі не супроводжується розщеплюванням лізосом, як при звичайному фагоцитозі. Цей процес управляється самими хламідіями, оскільки показано, що якщо в результаті фагоцитозу захоплюються хламідії, убиті нагріванням, то злиття лізосом не відбувається [Friis, 1972]. Неспеціалізовані клітини (не здібні до активного фагоцитозу) стимулюються хламідіями так, щоб вони їх захоплювали [Byrne, Moulder, 1978] [2].
Можна виділити 4 основних шляхи, по яких можуть реалізуватися подальші етапи взаємодії хламідій з клітиною-хазяїном (Шаткін А.А., Мавров І.І., 1983) [19]:
· Деструкція хламідії в фаголізосомній системі клітини-хазяїна
· Продуктивний цикл розвитку хламідій - репродукція
· L-подібна трансформація хламідій
· Персистенція хламідій.
Через 4-6 годин після зараження ЕТ [5], елементарні тільця розташовуються у вакуолі, оточеною деріватною мембраною, що захищає їх від дії лізоцимів. На відміну від вірусів хламідії не мають екліпсо-фази []. Деструкції, що далі не піддалися, в сприятливих умовах функціонування клітки-господаря, вступають в цикл розвитку, який протікає по відомій схемі: ЕТ реорганізуються у вегетативну форму - РT (через ПT); РT вступають в клітинний цикл, діляться бінарно (всього 8-12 циклів); дочірні РT перетворяться в ПT і далі в ЕТ- інфекційні форми нового покоління мікроорганізму. Цикл розвитку звичайно займає 48-72 години.
Вегетативну стадію в циклі розвитку хламідії, зокрема ділення і реорганізацію РT, можна розглядати як другий критичний етап взаємодії паразита і хазяїна. РT високочутливі до несприятливих екзогенних (наприклад, антибактеріальні препарати) і ендогенних дій, які можуть зумовити загибель паразита, що розмножується. В той же час антибіотики практично не діють на інфекційні ЕТ [20].
За несприятливих умов, на етапі функціонування РT, в цитоплазматичному включенні можуть бути виявлені аномальні форми хламідій з різними дефектами клітинної стінки. Ці утворення, морфологічно схожі з L-формами бактерій, закономірно утворюються під впливом тих, що традиційних трансформують агентів - Пеніциліну і ін. (Matsumoto A. et al., 1970). L-подібна трансформація хламідії може бути основою одного з механізмів, що індукують виникнення персистентної хламідійної інфекції, при безсимптомному перебігу яких спостерігається тривале перебування хламідій в клітині-хазяїна (Шаткін А.А., Попов В.Л., 1986).
Персистенція хламідій може спостерігатися при всіх відомих хламідіозах і у ряді випадків має надійне мікробіологічне підтвердження. Проте знання про цей шлях взаємодії паразита з хазяїном вкрай обмежені [5]. Схематично цей шлях з боку господаря може бути пов'язаний з нездатністю забезпечити відтворення паразита необхідними метаболітами і / або використанням обмежених механізмів захисту, здатних лише інгібірувати розмноження, але не елімінувати патогенний агент. Як і інші шляхи взаємодії, персистенція хламідій поза сумнівом контролюється імунною системою. Відомо, що лімфоцити від імунних тварин оберігають персистентно інфіковані макрофаги від активації хламідійної інфекції. Відома і активна течія і генералізує локалізованих експериментальних хламідійних інфекцій у іммуносупресованих тварин. Активний розвиток цього напряму досліджень є одним з актуальних завдань хламідіологів [2].
В процесі взаємодії з хламідією клітина-хазяїн піддається дії її чинників патогенності, що викликають різні порушення життєдіяльності клітки. При продуктивному циклі розвитку хламідій в клітині-хазяїна послідовно пригнічуються життєві функції, що приводять до її загибелі і руйнування. Цей результат, як правило, наступає в період завершення циклу розвитку хламідій. Суть подій, що протікають в клітині-хазяїна при L-подібній трансформації і персистенції хламідій, залишається нерозкритою. Функціонально в цих ситуаціях реалізується часовий збалансований стан, результат якого теоретично може бути на користь кожного із співчленів цієї системи.
Доля паразита і клітина-хазяїна при різних шляхах взаємодії в умовах природної інфекції багато в чому залежить від подій, що протікають на рівні макроорганізму. У цих умовах виявляються особливості паразитизму хламідій, характерні для внутріклітинних прокаріотів:
• Здатність індукувати фагоцитоз
• Протистояти деструктивній дії клітини-хазяїна
• Розмножуватися в клітині-хазяїна і тривалий час не викликати фатальних патологічних зміні
• У несприятливих умовах переживати або гинути під впливом клітини-хазяїна або екзогенних чинників [5].
Похожие работы

... а присутня тільки особлива система мембран, яка містить дві первинні дегідрогенази дихального ланцюга. Під дією кисню ці мембрани зливаються, утворюють складки і перетворюються в істинні мітохондрії, які містять цитохроми. РОЗДІЛ 2. МЕТОДИ ДОСЛІДЖЕННЯ МІТОХОНДРІЙ Значних успіхів було досягнуто у вивченні мітохондрій, при цьому значну роль зіграла електронна мікроскопія, яка дала можливість ...
... зі змістом білків 8 - 58 %, вуглеводів – 6 - 37 н жирів – 4 - 85 %. У залежності від умов вирощування в значній мірі змінюється також зміст вільних амінокислот, пігментів, вітамінів, мікроелементів. Водорості, особливо мікроскопічні, характеризуються найбільш високим КПД засвоєння світлової енергії в порівнянні з іншими фотосинтезуючими організмами. Багато видів здатні до ефективної утилізації сві ...
... синтезовані хімічним шляхом з'єднання, що не зустрічаються в природі, також атакуються мікробами. Всі викладені вище відомості, що становлять лише фрагменти того, що відомо сучасній мікробіології, та екології бактерії дають підставу зробити висновок про величезну й виняткову роль мікроорганізмів у круговороті речовин у природі. Види зелених водоростей та їх екологічне значення Зелені водорості ...
... Кролівництво та хутрове звірівництво кролі лисиця норка нутрія песець Рикетсійній кератокон'юктивіт Загальна інформація Рикетсійний кератокон’юнктивіт (keratokonynctivitis richetsion) – гостре інфекційне захворювання тварин з ураженням очей з розвитком катарального кон’юнктивіту і кератиту. Патогенез Не вивчено. Симптоми Інкубаційний період триває 10-12 діб. Перебіг хвороби гострий ...
0 комментариев