Навигация
Роль метилирования ДНК в контроле генома
33823
знака
0
таблиц
8
изображений
2. Роль метилирования ДНК в контроле генома
Одним из способов видоизменения гена является его замена на бессмысленную последовательность ДНК — тогда ген "выключается". Исследователи систематически "выключают" гены и наблюдают, к каким последствиям на уровне организма приводит это "выключение". Такая методика называется "генетический нокаут" (gene knockout).
Она позволяет подробно изучить функцию конкретного гена во время эмбрионального развития и после рождения животного. Можно проследить, как каждый ген влияет на развитие организма и возникновение той или иной патологии. В связи с этим, метод ещё получил название "генетическое планирование". К настоящему моменту уже проведены опыты по "выключению" десяти тысяч генов мыши, это половина всего мышиного генома.
Для развития организма достаточно одной клетки с единичной (конечно же, диплоидной) копией ДНК, которая при делении точно воспроизводится от клетки к клетке. Это относится практически ко всем живым многоклеточным существам. У человека все его клетки содержат идентичную ДНК. Клетки крови, печени, мозга, стволовые клетки - все они одинаковы по ДНК. Чем же определяется многообразие имеющихся у человека высокоспециализированных клеток и тканей? Это достигается за счет включения или выключения генов, ответственных за специализацию клетки. Именно механизмы контроля работы, или как принято говорить, экспрессии генов являются основной темой исследований нашей группы. Одним из таких механизмов является метилирование генов - ковалентное присоединение метильной группы в 5 положении пиримидинового кольца цитозина. ДНК, содержащая метилированные цитозины, является транскрипционно неактивной, и гены, располагающиеся вблизи метилированных районов, молчат.
Роль метилирования ДНК и механизм его негативного воздействия на работу генов в процессе жизнедеятельности позвоночных организмов (на моделях лягушки, мыши и клеточных линий человека).
Метилирование ДНК у позвоночных приобретает смысловую нагрузку в виде подавления транскрипции близлежащих генов двумя основными способами: (А) за счет прямого воздействия на ДНК, в составе которой метилированный цитозин ингибирует связывание транскрипционного фактора со своим участком, и (Б) за счет специфического связывания с метилированным районом специализированных метил-ДНК узнающих белков, которые, в свою очередь, привлекают сложные механизмы подавления транскрипции путем модификации близлежащих гистонов.
Как пример охарактеризован белок Каизо. Каизо, с одной стороны, связывается с катенином р120, а с другой - способен подавлять транскрипционную активность метилированных генов. Каизо имеет доменную структуру и состоит из N-концевого BTB/POZ домена и C-концевых цинковых пальцев типа C2H2. Цинковые пальцы специфично связывают 5-метил цитозин содержащую ДНК.
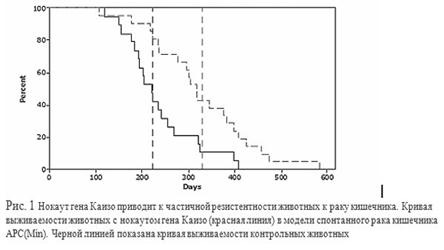
Нокаут гена Каизо приводит к частичной резистентности животных к раку кишечника. Кривая выживаемости животных с нокаутом гена Каизо (красная линия) в модели спонтанного рака кишечника APC(Min). Черной линией показана кривая выживаемости контрольных животных
Проведен генетический нокаут гена Каизо у мышей. Показано, что животные без Каизо (Каизо-КО) развиваются нормально и не имеют выраженных патологий . При переведении Каизо-КО животных на генетический фон с высоким процентом спонтанных опухолей кишечника происходит увеличение срока жизни животных, уменьшается средний размер полипов в кишечнике. Таким образом, установлено, что без Каизо происходит замедление роста опухолей кишечника в APC(Min) моделях Напротив, при выключении гена Каизо в зиготах лягушки происходит апоптотическая смерть клеток эмбрионов на стадии нейрулы. В отличие от мыши, ген Каизо является необходимым для жизнедеятельности земноводных. Эти данные были подтверждены и на рыбах Danio Rerio . Получены линии клеток с множественными генетическими нокаутами генов MBD2, MeCP2 и Каизо. Показано, что белки MBD2, MeCP2 и Каизо оказывают синергетическое действие (репрессируют) метилированный промотор гена Xist . Показано, что мутации в гене Каизо не являются ключевыми в инициировании синдрома Ретта (нейродегенративное заболевание у девочек) , хотя другой метил ДНК связывающий белок MeCP2 напрямую вовлечен в эту патологию.
В геноме позвоночных найдены и охарактеризованы два гена, кодирующих белки ZBTB4 и ZBTB38, которые по своей аминокислотной последовательности и расположению консервативных доменов являются родственниками Каизо. Показано, что эти два белка являются метил ДНК зависимыми транскрипционными репрессорами.
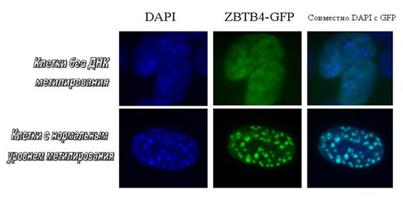
Каизо подобный белок ZBTB4 расположен в метилированных участках гетерохроматина. Клетки без метилирования - мышиные эмбриональные фибробласты с генетической делецией генов dnmt1-/- и p53-/-. Клетки с нормальным уровнем метилирования - мышиные эмбриональные фибробласты p53-/-. В случае dnmt1-/- p53-/- клеток видно отсутствие корреляции между локализацией ZBTB4 в гетерохроматине (DAPI), в то время, как в p53-/- клетках видна полная ко-локализация ZBTB4 и гетерохроматина.
3. "Программируемый нокаут генов"
Следует отметить, что не все гены можно инактивировать на стадии зародыша. И, естественно, нельзя получить клетки или животных, нокаутированных по так называемым генам домашнего хозяйства. Однако для генов, принимающих участие в эмбриональном развитии, разработан подход, позволяющий проводить их инактивацию после развития организма. Данная стратегия позволяет "выключать" гены в определенных условиях ("программируемый нокаут генов", англ. conditional knockout), а именно в необходимой исследователю ткани или группе клеток и/или под воздействием индуцирующего вещества.
Это можно осуществить с помощью методики, сочетающей гомологичную рекомбинацию, как в случае классического нокаута генов, и системы сайт-специфической рекомбинации. Сайт-специфические рекомбиназы – это ферменты, узнающие особые участки ДНК и совершающие обмен между ними. Наиболее часто используются Cre рекомбиназа бактериофага P1 и Flp рекомбиназа (флипаза) дрожжей. Эти ферменты распознают нуклеотидные последовательности в 34 основания, называемые, соответственно, loxP и frt сайты [16]. Если эти последовательности расположены в одной ориентации, рекомбинация по ним приведет к делеции фланкированного участка. Если же ориентация последовательностей различна, то это приведет к инверсии фрагмента между ними (рис. 5).
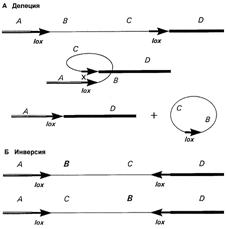
Рис. 5. Схема механизма сайт-специфической рекомбинации. В зависимости от ориентации loxP-сайтов происходит либо делеция, либо инверсия фланкированого фрагмента [20].
Использование стратегии "программируемого нокаута гена" требует создания двух линий мышей. Линия А несет интегрированную в геном последовательность гена Cre под контролем ткане-специфичного или индуцибельного промотора. Линия В содержит два loxP сайта, фланкирующих подлежащую удалению последовательность исследуемого гена (экзон, промотор и т.д.) [25]. Следует отметить, что вставки в геномную последовательность loxP сайтов и гена Cre осуществляются с использованием тех же приемов, что и при классическом нокауте генов, но не должны затрагивать функциональные последовательности (рис. 6).
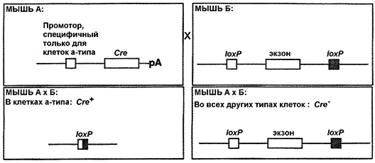
Рис. 6. Схема использования тканеспецифичной Cre-loxP рекомбинации для получения мышей с программируемым нокаутом гена [25].
Полученные таким образом гомозиготные линии мышей скрещивают. У потомков от этого скрещивания исследуемый ген будет инактивирован в ткани или группе клеток, где будет активен промотор, контролирующий активность гена Cre [25]. Используя стратегию "программируемой инактивации гена", можно добиться результатов, недоступных при использовании стандартной процедуры нокаута генов.
Похожие работы
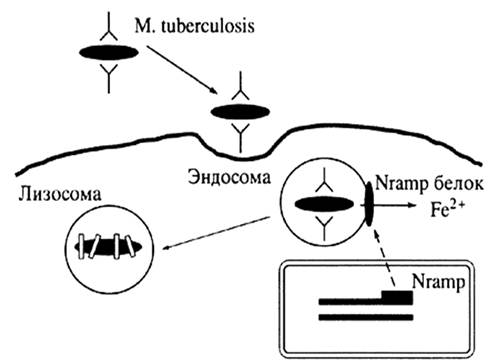
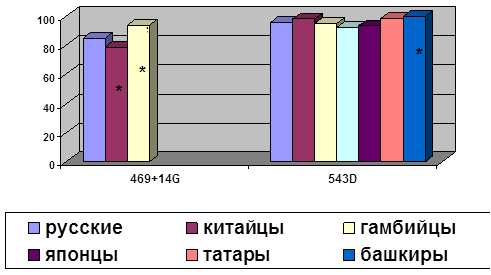
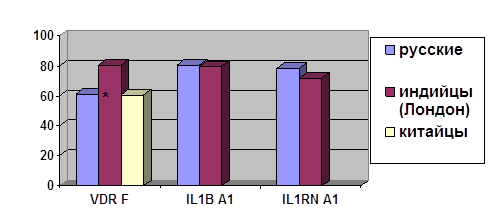
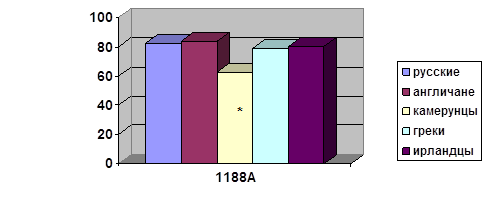
... о связи между частотой аллелей генов-кандидатов ТБ и особенностями распространения этого инфекционного заболевания. Выводы Выявлены особенности в распределении частот аллелей, изученных генов–кандидатов подвержености к туберкулезу, у русских жителей г. Томска по сравнению с другими популяциями мира. При сравнении с тувинцами показаны отличия в распределении генотипов и частот аллелей по всем ...
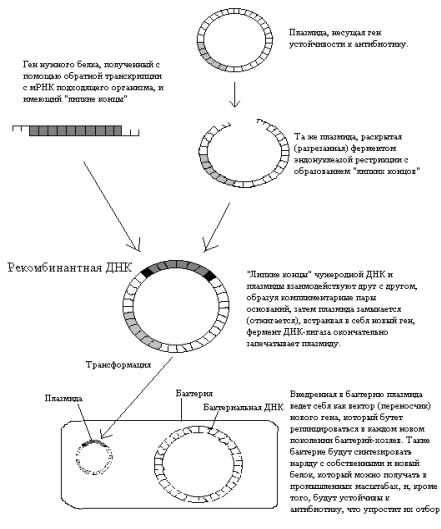
... в целях получения прибыли, к монополизации рынка продовольствия и не гарантирует безопасное и полезное для общества применение данной технологии. Краткая история возникновения генетически модифицированных организмов Истоки развития генной инженерии растений лежат в 1977 году, когда и произошло открытие, позволившее использовать почвенный микроорганизм Agrobacterium tumefaciens в качестве ...
... зеркальная копия этой молекулы, а потом копия копии, и возникает молекула, тождественная исходной. ДНК – это единственная саморазмножающаяся молекула в нашем организме. Какие же задачи будет решать биология в 21 веке ? В основном биологи считают, что первая задача это старение. Здесь две точки зрения. Одна – общепринятая, что старение – это поломка сложной системы, которая рано или поздно ...
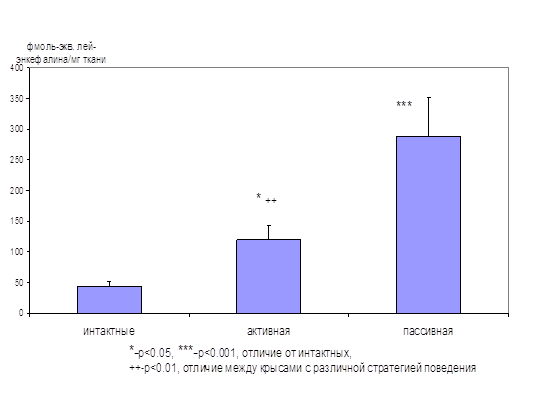

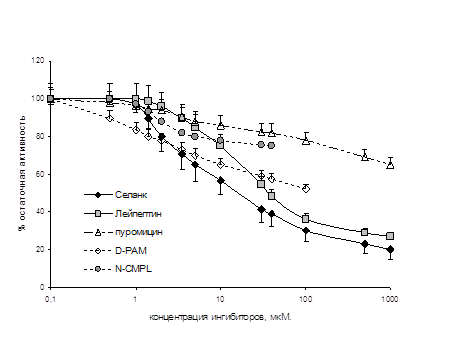
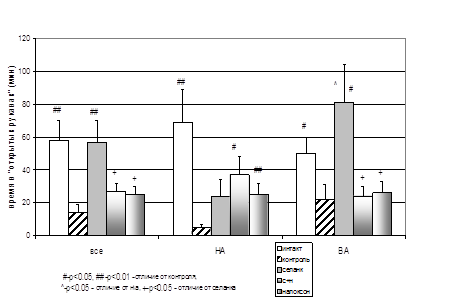
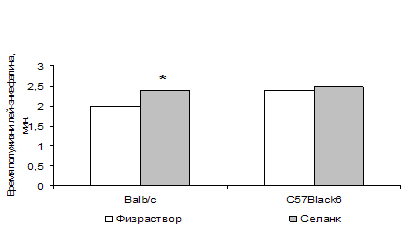
... энкефалиназ при ГТР. То есть, истощение эндогенной опиоидной системы, выражающееся в недостатке эндогенных ингибиторов ЭДФ, по-видимому, является одним из механизмов стресс-индуцированных нарушений, существенных в патогенезе некоторых видов тревожных расстройств. Полученные в нашем исследовании данные позволяют предполагать, что ГТР является одной из таких нозологий. То есть стимуляция опиоидной ...
0 комментариев