Навигация
Строение и состав живой клетки
17623
знака
0
таблиц
6
изображений
Введение
Строение и состав живой клетки
Введение
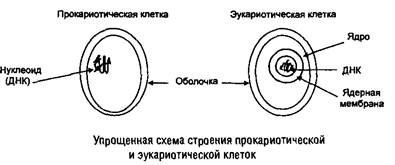
Все живые организмы состоят из клеток и подразделяются на одноклеточные и многоклеточные. Популяция одноклеточных организмов включает клетки одного типа или небольшого количества типов, тогда как у многоклеточных организмов клетки специализированы. По типу строения клетки подразделяются на прокариотческие и эукариотические.
Все клеточные организмы имеют сходный химический состав и содержат три основных типа макромолекул: ДНК, PHK белки, а также полисахаридные и липидные компоненты. Кроме того, в клетках присутствуют переменные количества низкомолекулярных веществ — субстратов и продуктов энергетических и конструктивных процессов (аминокислоты, сахара, нуклеотиды и др.). В среднем около 80% клеточной массы составляет вода.
1. Клеточные стенки и клеточные мембраны
Как правило, клетки окружены двумя оболочками: клеточной стенкой и цитоплазматической мембраной, для эукариотических клеток применяют название плазмолемма.
Клеточная стенка обеспечивает механическую прочность клетки, придавая ей жесткую (ригидную) структуру, благодаря чему клетка выдерживает высокое внутреннее осмотическое давление (5—20 МПа). Кроме того, клеточная стенка может обусловливать некоторую степень избирательной проницаемости для низкомолекулярных веществ, а также способность взаимодействовать с другими клетками, вирусами и физическими поверхностями. Строение клеточной стенки у разных организмов имеет свои особенности.
Клетки большинства тканей многоклеточных животных не содержат выраженной клеточной стенки. Растительные клетки, напротив, имеют очень сложную клеточную стенку, построенную из целлюлозных микрофибрилл, погруженных в матрикс (из пектина и гемицеллюлоз).
Клеточные стенки дрожжей и мицелиальных грибов состоят из гомо- и гетерополисахаридов (глюканов, хитина) и белкового комплекса, выполняющего антигенную роль. Толщина этих слоев достигает 1 мкм.
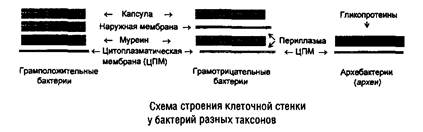
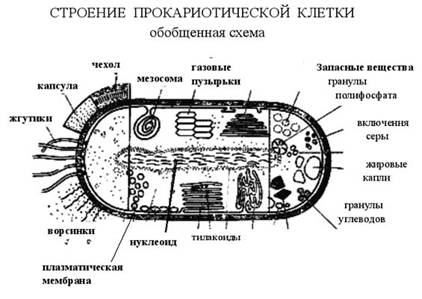
Особенности клеточных стенок бактерий связаны со способностью бактерий окрашиваться по Граму. Клеточная стенка громположительных бактерий построена в основном из гетерополисахарида муреина (пептидогликана), содержащего аминокислотные «мостики». В состав клеточной стенки входят также тейхоевые кислоты (рибит- или глицеринтейхоевые), ковалентно связанные с мурамовой кислотой пептидогликана. У грамотрицательных бактерий слой муреина невелик, но в клеточной стенке присутствует наружная мембрана, построенная из фосфолипидов, белков и липопо-лисахарида, обеспечивающая некоторую степень избирательной проницаемости и содержащая рецепторы фагов и антигены. Толщина клеточной стенки составляет от 15 до 80 нм. Существуют бактерии, полностью лишенные клеточной стенки (микоплазмы). У представителей архебактерий (архей) в клеточной стенке отсутствует муреин (иногда содержится отличающийся по составу псевдомуреин), а осмопротекторную роль выполняет слой гликопротеинов (рис. 3).
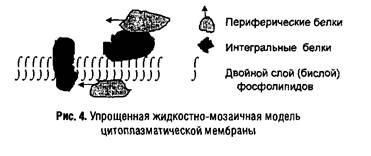
Цитоплазматическая мембрана состоит из белков и липидов в соотношении от 1: 4 (в миелине, мембране нервных волокон) до 4:1 (в мембранах бактерий). Во многих мембранах присутствуют небольшие количества углеводов (до 5%) и следы РНК, а также неорганические катионы (в основном Са2+ и Mg2+). Существенную часть мембран составляет вода — она участвует в формировании гидрофобных (энтропийных) связей между компонентами мембраны. Еще в 1930 г. Д. Даниэлян предложил модель мембраны в виде двойного липидного слоя или бислоя. Развитием этих представлений является жидкостно-мозаичная модель, предложенная С.Д. Сингером и Г. Николсоном в 1972 г., согласно которой мембранные белки подразделяются на периферические (слабо связанные с мембраной) и интегральные (локализованные в липидном бислое). Последние свободно плавают в «липидном море», перемещаясь в латеральном направлении (в плоскости мембраны), но неспособны «пронырнуть» на противоположную ее сторону. Таким образом, мембраны оказываются асимметричными, по крайней мере, в отношении белкового состава. Асимметрия наблюдается и в расположении липидов в мембране, но менее выражена. Некоторые из интегральных белков имеют трансмембранную ориентацию и выполняют функции переносчиков субстратов или сигналов. Схема подобной мембраны представлена на рис. 4.
2. Состав мембранных липидов
Основная часть мембранных липидов представлена фосфолипидами, в основе которых лежит глицерин-3-фосфат. Такие фосфолипиды называют фосфоацилгпицеринами. В их молекуле гидроксильные группы глицерина при Cf и С2 этерифицированы жирными кислотами, а остаток фосфорной кислоты либо остается свободным, как в диацилглицерин-3-фосфате (или фосфатидной кислоте), либо этерифицирован спиртовыми гидроксилами серина, этаноламина, холина, инозита, глицерина или глицерина и лизина. Соответственно они называются: фосфатидилсерин, фосфатидилэтаноламин (кефалин), фосфатидилхолин (лецитин), фосфатидилинозит, фосфатидилглицерин, лизилфосфатидилглицерин. Объединение молекул фосфатидной кислоты и фосфатидилглицерина дает дифосфатидилглицерин (кардиолипин), обладающий иммунологическими свойствами (рис. 5).

Удаление одной из ацильных групп приводит к лизофосфолипидам, которые обладают выраженными поверхностно-активными свойствами и могут, например, способствовать эмульгированию жиров в кишечнике.
Мембраны грамположительных бактерий содержат все эти фосфолипиды (кроме лецитина), тогда как у грамотрицательных бактерий основным, а то и единственным мембранным фосфолипидом является фосфатидилэтаноламин.
Соотношение различных классов фосфолипидов в мембране существенно зависит от условий выращивания организма. Например, при снижении рН среды в мембранах бактерий начинают преобладать положительно заряженные фосфолипиды (аминоацилфосфатидилглицерины, фосфатидилэтаноламин), наличие которых существенно уменьшает проницаемость мембраны для протонов. В большинстве случаев фосфолипиды представляют собой сложные эфиры глицерина или других многоатомных спиртов, но существуют липиды, в которых одна из спиртовых групп глицерина образует простую эфирную связь с ал килами или алкенами. Необычные липиды найдены у большой группы прокариот, так называемых архебактерий (или архей), в качестве простых эфиров глицерина и длинноцепочечных С-20, С-40-гидроизопреноидных спиртов (полипренолов), точнее, дифитанилдиглицериновых диэфиров или дибифитанилдиглицериновых тетраэфиров (рис. 6).
Мембрана, образованная тетраэфирами, уже не может рассматриваться как двойной липидный слой в истинном значении этого понятия и не подвергается расщеплению по гидрофобной сердцевине методом «замораживания-скалывания» (freeze-fracture). Она более устойчива к стрессовым воздействиям окружающей среды, что позволяет микроорганизмам, имеющим такие мембраны, существовать в экстремальных условиях рН и температуры (экстремофилы). У термофилов дополнительная стабилизация достигается формированием в полипренольной цепи пятичленньгх циклов, уменьшающих вращательную подвижность углеводородных цепей.

Кроме фосфолипидов, построенных на основе глицерина, в клетках эу- и прокариот встречаются фосфолипиды, которые являются производными диодов: этиленгпиколя, 1,2- и 1,3-пропандиолов, 1,3-, 1,4- и 2,3-бутандиолов, а также 1,5-пентандиола. Среди диольных липидов встречаются моно- и диацильные производные — это сложные эфиры различных жирных кислот; простые эфиры, смешанные алкильные (или алкенильные) и ацильные производные, диольные аналоги фосфатидилхолина, фосфатидилэтаноламина и др. Обычно содержание диольных липидов составляет только 0,5—1,5% содержания глицериновых липидов. Но некоторые морские моллюски и иглокожие содержат в тканях примерно равные количества производных глицерина и этиленгликоля. Содержание диольных липидов уменьшается в течение зимы, поэтому они предположительно могут использоваться как запасные вещества. Замечено также их увеличение в процессе регенерации печени у крыс.
Другим классом «безглицериновых» липидов являются сфинголипиды, основу которых составляет алифатический аминоспирт сфингозин (или дигидросфингозин) (рис. 7). Построенные на его основе церамиды широко распространены в тканях растений и животных, но количество их незначительно. Они обнаружены также в пилях Escherichia coli. Сфингомиелины входят в состав нервной ткани, липидов крови и некоторых других компонентов клеток животных.

В большинстве биологических мембран содержатся также гликолипиды. В клетках животных они, как правило, являются производными сфингозина, у которого к первичному гидроксилу присоединен остаток сахара или олигосахарида. Если сахарами являются глюкоза или галактоза, такие липиды называются цереброзидами (особенно много их в тканях мозга). Если углеводной частью является олигосахарид, речь идет о ганглиозидах (выделены из ганглий, мозга и ряда других тканей). Ганглиозиды найдены также в пилях некоторых бактерий (Neisseria) и определяют прилипание этих патогенов к поверхности клеток животного организма. У прокариот (в основном у грамположительных бактерий и цианобактерий) гликолипиды также содержат глюкозу, галактозу и маннозу. Их количество в мембранах невелико, но при лимитировании фосфором может возрастать, при этом гликолипиды, по-видимому, замещают фосфолипиды. Наконец, в мембранах большинства эукариот (а также у Mycoplasma) содержатся стероиды, в основном холестерин (у животных), эргостерин (у дрожжей) и стигмастерин (у растений). Микоплазмы неспособны синтезировать стерины, но требуют присутствия их в среде для стабилизации клеточной мембраны.
Похожие работы
... веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза. генетика клетка онтогенез гибрид Законы Г. Менделя В своих опытах по скрещиванию Мендель применял гибридологический метод. Используя этот метод, он изучал наследование по отдельным признакам, а не по всему комплексу, ...
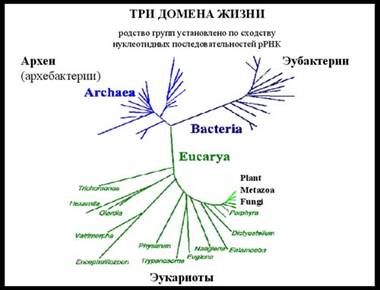

... » обозначает группу (рангом выше царства), объединяющую разные организмы, обладающие определенным набором общих черт. Что же общего у бактерий и архей, что отличает их от эукариот? Строение клетки прокариот У прокариотических клеток есть цитоплазматическая мембрана, также как и эукариотических. У бактерий мембрана двуслойная (липидный бислой), у архей мембрана довольно часто бывает однослойной. ...

... России страдает от недостатка в питании витаминов и других функциональных веществ. Все большее распространение получают различные заболевания, снижается средняя продолжительность жизни. Химический состав плодов и овощей отличается большим разнообразием, он определяет цвет, вкус, запах, пищевую ценность продукта, сохраняемость и функциональные свойства. Энергетическая ценность большинства видов ...
кратце рассмотреть некоторые представления о биохимии живой клетки, как они сложились к началу 50-х годов прошлого столетия прочную «третичную структуру» фермента, так и расположение каталитического, или, как говорят, «активного» центра на поверхности его глобулы. Наконец, что же может представлять из себя сам активный центр? Да ничего иного, кроме строго фиксированного пространственного ...
0 комментариев