Навигация
Природа и состав жирных кислот в мембранных липидах
17623
знака
0
таблиц
6
изображений
3. Природа и состав жирных кислот в мембранных липидах
Природа жирных кислот в липидах мембран зависит как от вида организма, так и от условий его существования. Наиболее часто встречающиеся жирные кислоты липидов животных, растительных и прокариотных клеток.
С увеличением числа двойных связей значительно снижается температура плавления жирных кислот (а также содержащих эти кислоты липидов) и повышается их растворимость в неполярных растворителях. Поскольку функциональная активность мембранных белков регулируется фазовым состоянием липидов мембраны (как правило, в жидком состоянии их активность выше), при снижении температуры в мембране должно повышаться содержание ненасыщенных кислот. Благодаря постоянству внутренних условий (гомеостазу) животного макроорганизма влияние температуры на жирнокислотный состав липидов обычно проявляется слабо, но, например, в липидах нижних конечностей пингвинов повышено содержание ненасыщенных жирных кислот.
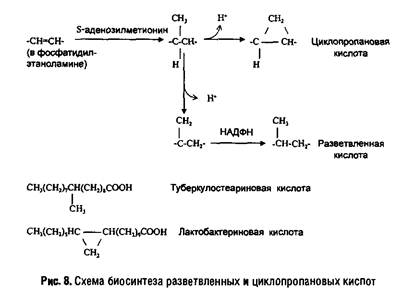
В мембранах прокариот разнообразие жирных кислот довольно велико. Особенностью бактерий является наличие разветвленных и циклопропановых кислот. У некоторых грамположительных бактерий (например, Micrococcus luteus) 90% жирных кислот липидов составляют разветвленные кислоты, а ненасыщенные кислоты практически отсутствуют. Грамотрицательные бактерии содержат смесь насыщенных и ненасыщенных кислот с преобладанием С-16 и С-18 кислот, а также циклопропановые кислоты (например, лактобацилловую кислоту). Пути их биосинтеза показаны на рис. 8.
Изменение температуры существенно влияет на соотношение насыщенных и ненасыщенных жирных кислот в мембранах прокариот, при этом снижение температуры культивирования приводит к увеличению доли ненасыщенных кислот. Предельными случаями являются психрофилы, растущие при температурах, близких к 0"С, у которых присутствуют практически только ненасыщенные жирные кислоты, а также термофилы (привычная температурная среда выше 60°С) — у них все жирные кислоты липидов насыщенные.
Тем не менее состав жирных кислот фосфолипидов у бактерий при культивировании в стандартных условиях достаточно постоянен, и его можно использовать как таксономический признак — в интересах классификации или при анализе состава микробных сообществ без изолирования в чистых культурах отдельных представителей.
Для изучения структуры, состава и функции органелл необходимо, как правило, изолировать их из клетки в чистом виде. Обычно это достигается методом дифференциального центрифугирования после разрушения клеток в механических или ультразвуковых гомогенизаторах.
Кратко охарактеризуем основные процессы и органеллы (в алфавитном порядке).
Аэросомы — однослойные везикулы, мембрана которых построена только из белка. Способствуют повышению плавучести клеток, так как в них содержится газовая фаза, совпадающая по составу с газовой фазой окружающей среды.
Вакуоли — мембранные образования, служащие для поддержания тургорного давления, запасания различных веществ, а также выполняющие лизосомные функции.
Внехромосомная ДНК. В митохондриях и хлоропластах содержится ДНК, образующая нуклеоид бактериального типа. Заключенная в ней генетическая информация не дублируется в ядерной ДНК и способна к автономному выражению в белках посредством собственных систем транскрипции и трансляции (включающих рибосомы 70 S бактериального типа).
У прокариот внехромосомная ДНК организована в виде плазмид, которые могут существовать и реплицироваться автономно или в интегрированном в хромосому состоянии (например, в виде профага).
Гидрогеносомы — окружены однослойной мембраной и содержат комплекс пируватдегидрогеназ (у трихомонад).
Гистоны — представляют собой положительно заряженные (основные) белки, входящие в состав хромосом в комплексе с ДНК (обнаружены также у архебактерий).
Гликосомы — окружены однослойной мембраной и содержат ферменты гликолиза (у некоторых протозойных микроорганизмов, в частности у возбудителей сонной болезни).
Глиоксисомы — разновидность пероксисом — место локализации ферментов глиоксалатного шунта, участвующих в превращении запасных жиров в углеводы. Поэтому они тесно ассоциированы со сферосомами, жирозапасающими органеллами растительных клеток.
Жгутики и реснички представляют собой аппарат, определяющий подвижность клеток или их способность создавать поток окружающей среды к органам поглощения пищи. Жгутики эукариот и прокариот сильно различаются по строению и составу.
Карбоксисомы — окружены однослойной мембраной и содержат ключевые ферменты фиксации углекислоты в цикле Кальвина (у фототрофных и некоторых хемолитотрофных прокариот).
Лизосомы — мембранные везикулы, содержащие гидролитические ферменты, участвующие в круговороте белков, полисахаридов, липидов и нуклеиновых кислот. Для предотвращения неорганизованного действия этих ферментов они заключены в органеллу, окруженную однослойной мембраной. Некоторые наследственные болезни связаны с недостаточностью лизосомных ферментов.
Микротрубочки и микрофиламенты, по-видимому, исполняют роль цитоскелета и формируются из белка тубулина. Они входят в состав центриолей, играющих важную роль в делении ядра, а также в состав жгутиков и ресничек.
Митоз — деление ядра, сопровождающееся удвоением числа хромосом; мейоз — деление ядра без удвоения числа хромосом, в результате чего образуются гаплоидные клетки.
Митохондрии являются местом осуществления окислительных процессов, в них локализованы ферменты цикла трикарбоновых кислот, дыхательная цепь, система окислительного фосфорили-рования. В клетках содержится от 1 до нескольких тысяч митохондрий. Митохондрии содержат внехромосомную ДНК, способны к самовоспроизведению и, возможно, ведут свое начало от прока-риотических клеток (эндосимбиотическая гипотеза). Однако часть белков этих органелл кодируется ядерными генами, поэтому они не культивируются автономно. У прокариот сходные по функциям структуры, происходящие из цитоплазматической мембраны, называют мезо сомами.
Пероксисомы — органеллы, окруженные однослойной мембраной и содержащие ряд окислительных ферментов, а также каталазу, разрушающую перекись водорода, образующуюся в процессе окисления. Могут рассматриваться как более древние аналоги митохондрий и играют важную роль в обмене липидов.
Пили, фимбрии представляют собой выросты поверхности клеток и бывают простые и половые. Простые пили построены из белка пилина и играют важную роль в прикреплении бактерий к субстрату. На поверхности клетки их может быть 50—400 штук. Половые пили (1 на клетку) используются в процессе конъюгации. Выросты клеток, содержащие цитоплазму, называют простеками.
Они служат для увеличения поверхности клеток, прикрепления к субстрату или участвуют в почковании клеток.
Хитосомы — место локализации хитинсинтетазы, встречаются у грибов. Осуществляют функцию переноса микрофибрилл хитина к клеточной стенке.
Хлоропласты содержат хлорофилл и осуществляют процесс фотосинтеза у растений. Как и в митохондриях, в них содержится собственная ДНК. Аналогичные по функциям бактериальные структуры называют хроматофорами, а также тилакоидами и хлоросомами. В отличие от хлоропластов они не содержат ДНК и происходят из цитоплазматической мембраны.
Целлюлосомы — содержат комплекс ферментов и липидов. Определяют присоединение бактерий к целлюлозным субстратам и их расщепление.
Эндоплазматтеский ритикулум — это внутриклеточная сеть взаимосвязанных, ограниченных мембраной трубочек и пузырьков. Мембрана эндоплазматического ретикулума образует единое целое с ядерной мембраной. Если на наружной поверхности эндоплазматического ретикулума адсорбированы рибосомы, его называют шероховатым, если нет — гладким. Шероховатый эндоплазматический ритикулум — место синтеза секретируемых белков, которые попадают внутрь его каналов, а затем поступают в «цистерны» аппарата Гольджи (диктиосомы у растений), откуда переносятся внутри мембранных везикул либо в плазмалемму, либо в мембрану вакуолей (тонопласт) и секретируются внутрь вакуолей или в окружающую среду (экзоцитоз).
Ядро (нуклеоплазма) эукариотических клеток окружено двойной мембраной (плазмолеммой), содержащей поры (поросомы). В ядре присутствует ядрышко (место синтеза РНК и сборки рибосом). В отличие от ядра нуклеоид прокариотических клеток не отделен мембраной от цитоплазмы и представляет собой комплекс ДНК, РНК и белков.
Похожие работы
... веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза. генетика клетка онтогенез гибрид Законы Г. Менделя В своих опытах по скрещиванию Мендель применял гибридологический метод. Используя этот метод, он изучал наследование по отдельным признакам, а не по всему комплексу, ...

... » обозначает группу (рангом выше царства), объединяющую разные организмы, обладающие определенным набором общих черт. Что же общего у бактерий и архей, что отличает их от эукариот? Строение клетки прокариот У прокариотических клеток есть цитоплазматическая мембрана, также как и эукариотических. У бактерий мембрана двуслойная (липидный бислой), у архей мембрана довольно часто бывает однослойной. ...

... России страдает от недостатка в питании витаминов и других функциональных веществ. Все большее распространение получают различные заболевания, снижается средняя продолжительность жизни. Химический состав плодов и овощей отличается большим разнообразием, он определяет цвет, вкус, запах, пищевую ценность продукта, сохраняемость и функциональные свойства. Энергетическая ценность большинства видов ...
кратце рассмотреть некоторые представления о биохимии живой клетки, как они сложились к началу 50-х годов прошлого столетия прочную «третичную структуру» фермента, так и расположение каталитического, или, как говорят, «активного» центра на поверхности его глобулы. Наконец, что же может представлять из себя сам активный центр? Да ничего иного, кроме строго фиксированного пространственного ...
0 комментариев