Навигация
2. Водно-липидные смеси
Смеси липидов с водой отличаются выраженным полиморфизмом. Даже индивидуальные очищенные липиды в гидратированном состоянии могут находиться в нескольких структурных модификациях. Какая из структур преобладает, зависит от таких параметров, как концентрация липида, температура, давление, ионная сила и рН. Особенно полезным при изучении типов структурной организации водно-липидных систем оказался метод дифракции рентгеновских лучей. При этом чаще всего варьируют концентрацию липида и температуру, а полученные данные представляют в виде фазовой диаграммы, показывающей, какую структуру система имеет в различных областях диаграммы «температура — концентрация». Наряду с дифракцией рентгеновских лучей для определения фазовых границ водно-липидных систем часто используют дифференциальную сканирующую калориметрию. Эти исследования проводят обычно при высоких концентрациях липида, однако многие структуры, обнаруженные при таких условиях, образуются также в липидных дисперсиях при большом избытке воды.
Основные типы структурной организации водно-липидных систем схематично представлены на рис. 2.4.
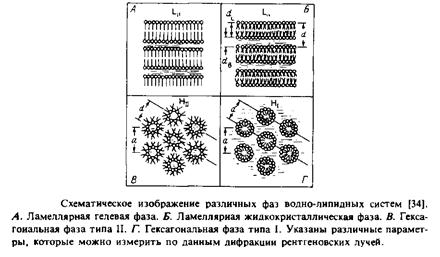
1.Ламеллярная жидкокристаллическая фаза, Считают, что именно в этой фазе находится основная масса липидов в биологических мембранах. Как свидетельствуют данные дифракции рентгеновских лучей, для этой фазы характерно упорядоченное расположение слоистых структур при значительной неупорядоченности ацильных цепей.
2.Ламеллярная гелевая фаза. Она образуется при низкой температуре теми липидами, которые формируют слоистые структуры. В этой фазе молекулы упакованы более плотно, а ацильные цепи намного более упорядочены и находятся преимущественно в полнос-тью-трянс-конфигурации, как и в липидных кристаллах. Поскольку цепи максимально вытянуты, толщина бислоя в фазе геля выше, чем в жидкокристаллической фазе. Плотность фазы геля несколько выше плотности жидкокристаллической фазы. В случае липидов, имеющих объемные полярные головки, ацильные цепи наклонены относительно поверхности бислоя подобно тому, как это наблюдается в некоторых липидных кристаллах. Наклон цепей обычно обозначают штрихом Интересно, что дисперсии фосфатидилхолина в растворах, содержащих некоторые спирты, в том числе и глицерол, образуют необычную фазу геля, в которой противолежащие половины «бислоя» своими ациль-ными цепями полностью проникают друг в друга. Биологическая роль этого явления неясна.
3. Гексагональная фаза I. В этом случае липидные молеку-
лы формируют цилиндрические структуры, поверхность которых образована полярными головками и контактирует с водой. Сами цилиндры упаковываются с образованием гексагональной решетки.
4. Гексагональная фаза II. Липиды также образуют цилиндры, но в этом случае полярные группы обращены внутрь цилиндра и формируют водный канал. Упаковка самих цилиндров также является гексагональной.
Очень важно, что некоторые липиды образуют небислойные структуры. Действительно, многие очищенные мембранные липиды не образуют стабильных бислоев, а предпочитают находится в гек-
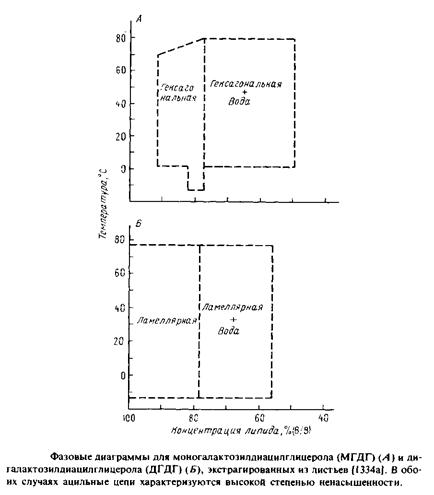
сагональной фазе Ни. В качестве примера можно упомянуть ненасыщенные фосфатидилэтаноламины, а также такой гликолипид, как моногалактозилдиацилглицерол. Причины такого поведения и его возможная биологическая значимость обсуждаются в следующих разделах.
2.1 ГИДРАТАЦИЯ ЛИПИДОВ
Параметры можно определить по данным дифракции рентгеновских лучей. Как правило, те параметры, которые зависят главным образом от длины ацильных цепей, почти не меняются при увеличении содержания воды в системе. Гидратация липидов происходит в результате связывания воды с полярными головками. Процесс гидратации активно изучали методами 'Н- и 2Н-ЯМР. Результаты, полученные при изучении фос-фатидилглицерола, фосфатидилхолина и фосфатидилэтаноламина методом 2Н-ЯМР, говорят о наличии гидратной оболочки из 11 — 16 молекул воды на одну молекулу липида. Эти молекулы быстро обмениваются с молекулами воды, находящимися в основном объеме. Согласно другим измерениям, полярная головка фосфатидилэтаноламина связывает меньше воды, чем полярная головка фосфатидилхолина. Было высказано предположение, что в случае ненасыщенных фосфатидилэтаноламинов слабая гидратация благоприятствует образованию неламеллярной гексагональной фазы Ни.
Многие липиды набухают в воде. Липиды, которые не несут заряда или являются в целом электрически нейтральными, не набухают совсем или набухают лишь в ограниченной степени до предельной толщины водной прослойки между ламеллами. При избытке воды сосуществуют две фазы — мультиламеллярная липидная фаза и вода, находящаяся в основном объеме. Заряженные липиды склонны к неограниченному набуханию и могут включать воду между ламеллами вплоть до пороговой точки, когда образуются две фазы — полностью гидратированные моноламелярные везикулы, находящиеся в равновесии с водой в основном объеме. Степень набухания и относительная стабильность мульти- и моноламеллярных структур определяются электростатическими взаимодействиями. При низкой ионной силе происходит дестабилизация мультиламеллярных структур. Неограниченное набухание может происходить и в том случае, когда в смеси липидов содержится всего несколько процентов заряженных липидов.
Поляризация молекул воды вблизи полярных липидных головок приводит к сильному отталкиванию при сближении двух бислоев. Эта «гидратационная сила» удерживает гидратированные бислои на расстоянии не менее 30 А друг от друга. Именно она создает основной энергетический барьер, который следует преодолеть, пытаясь осуществить слияние мембран. Возможно, фосфатидилэтаноламиновые везикулы склонны к агрегации именно потому, что степень гидратации их полярных головок относительно низка.
Исследования, проведенные методом ЭПР с помощью спиновых меток, способных реагировать на полярность своего окружения на различной глубине от поверхности бислоя, показывают, что вода частично проникает в углеводородную область бислоя, находящуюся в жидкокристаллическом состоянии. По данным нейтронного рассеяния, когда бислой находится в состоянии геля, вода не проникает глубже глицеролового остова липидных молекул.
Похожие работы
... является курсом, для изучения которого необходимо наличие знаний об основных принципах организации биологических молекул, строении и механизмах действия ферментов. Дисциплина биохимия мембран относится к дисциплинам специализации федерального компонента. 5. Распределение времени, отведенного на изучение дисциплины по учебному плану Форма учебной работы Форма обучения Очная По ...
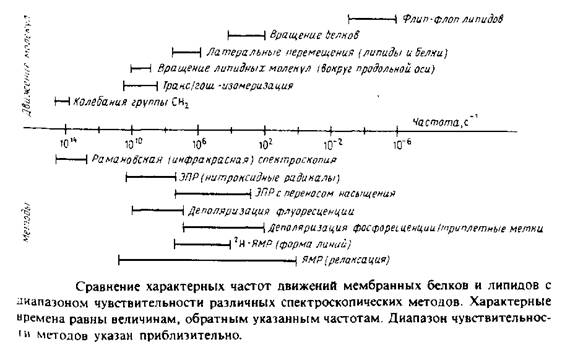
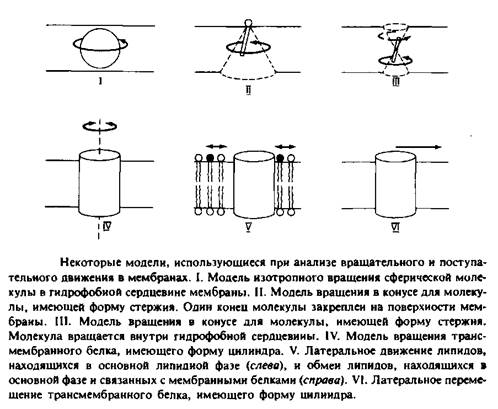
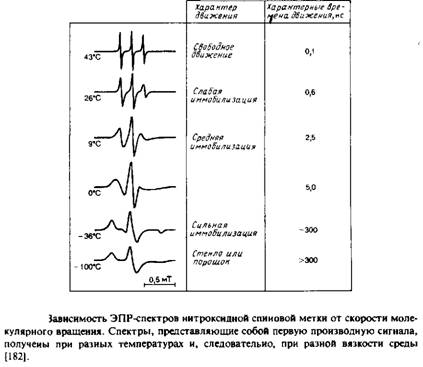
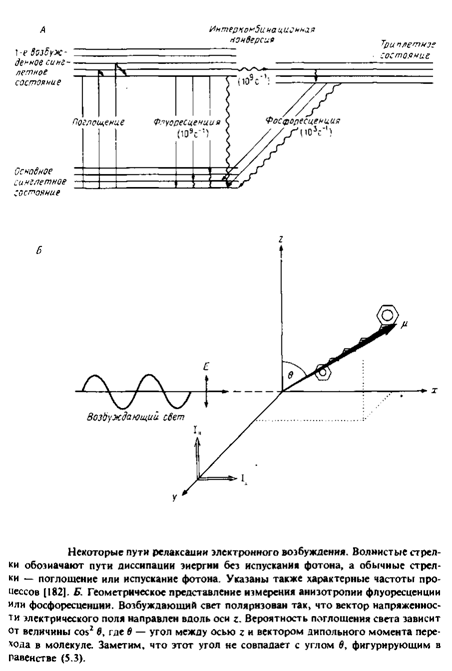
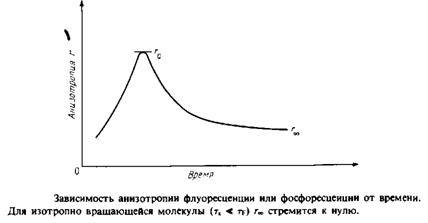
... к уменьшению латеральной диффузии. 5. Липидно-белковые взаимодействия Большинство методов, применяемых для изучения упорядоченности и динамических свойств мембран, используется и для исследования липидно-белковых взаимодействий. Работы по изучению этих взаимодействий были в основном направлены на выяснение влияния мембранных белков на физическое состояние липидов. Рассмотрим типичную мембрану ...

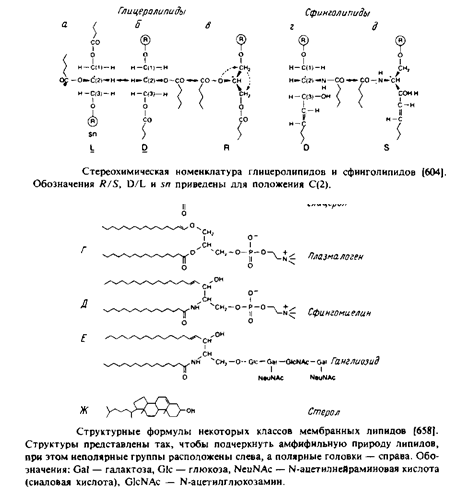
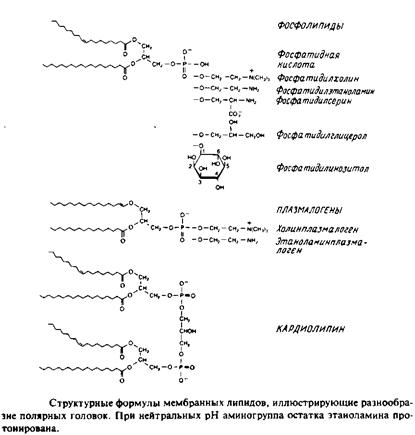


... животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы. Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. ...
... - восстановительные. 5.Ферменты патогенности (вирулентности). Ферментный состав клетки определяется геномом и является достаточно постоянным признаком. Знание биохимических свойств микроорганизмов позволяет идентифицировать их по набору ферментов. Основные продукты ферментирования углеводов и белков- кислота, газ, индол, сероводород, хотя реальный спектр для различных микроорганизмов намного ...
0 комментариев