Навигация
ОРИЕНТАЦИЯ ПОЛЯРНЫХ ГОЛОВОК ЛИПИДОВ В БИСЛОЕ
31746
знаков
0
таблиц
14
изображений
2.4. ОРИЕНТАЦИЯ ПОЛЯРНЫХ ГОЛОВОК ЛИПИДОВ В БИСЛОЕ
Данные ряда методов свидетельствуют о том, что в ламелляр-ных водно-фосфолипидных дисперсиях, как и в липидных кристаллах, полярные головки липидов в целом ориентированы параллельно плоскости бислоя. В случае фосфатидилхолинов такая ориентация присуща как гелевой, так и жидкокристаллической фазам, судя по результатам исследований методами дифракции нейтронов и рентгеновских лучей. Об этом же свидетельствуют и данные, полученные методом 2Н-ЯМР, хотя при этом не исключаются и другие интерпретации. Имеющиеся данные указывают на то, что пространственное расположение полярных головок фосфа-тидилглицерина, сфингомиелина и фосфатидилсерина сходно. Исследования методом 2Н-ЯМР интактных фибробластов мыши и выделенных из них мембран также показали, что полярные головки как фосфатидилхолина, так и фосфатидилэтаноламина ориентированы параллельно поверхности мембран. Однако по данным дифракции нейтронов у фосфатидилглицерола, выделенного из Е. coli, полярная головка ориентирована примерно под углом 30° к поверхности мембраны, что облегчает связывание отрицательно заряженных фосфатных групп с катионами.
На ориентацию и динамику полярных головок липидных молекул может влиять образование межмолекулярных водородных связей на поверхности мембраны. Донорами и акцепторами при Образовании этих связей могут служить такие липиды, как фосфатидилсерин, фосфатидилэтаноламин и различные гликолипиды. Исследования, проведенные на модельных мембранных системах, показывают, что водородные связи между полярными головками сохраняются даже в условиях гидратации мембранной поверхности, однако пока неизвестно, как образование этих связей может сказаться на структуре биологических мембран.
2.5. КОНФИГУРАЦИЯ И УПАКОВКА АЦИЛЬНЫХ ЦЕПЕЙ В БИСЛОЕ
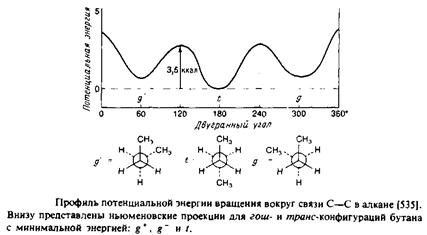
Рассмотрим сначала насыщенные углеводородные цепи. В них возможно свободное вращение вокруг каждой С—С-связи, характеризующееся энергетическим минимумом, особенно четким в случае ньюменовской проекции. Наиболее стабильна транс-конфигурация, при этом высота энергетического барьера для перехода через заслоненную конфигурацию в гош-форму составляет по оценкам 3,5 ккал/моль. В полностью m/wwc-конфигурации цепь максимально вытянута и не меняет своего направления, тогда как в гош-конформации ее направление меняется. Последовательность гош-транс-гош для трех смежных С—С-связей приводит к появлению в цепи излома, в результате чего участки цепи выше и ниже места излома оказываются значительно смещенными друг относительно друга. /ош-конфигурация в зависимости от направления вращения при переходе от Q к С4 обозначается как g+ или g". Кинки типа g + tg~ или g~ tg+ приводят к минимальному сдвигу цепи. Почти все двойные связи в мембранных липидах находятся в J/ис-конфигурации. Как и в случае гош-конфигурации, это приводит к изменению общего направления цепи. Наличие в углеводородных цепях кинков, двойных ^ыс-связей,
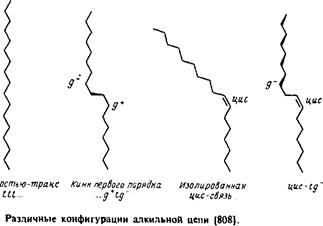
циклопропановых групп и других особенностей приводит к увеличению площади поперечного сечения цепи; это может иметь важные последствия для упаковки липидов в бислое. При этом стерические требования к упаковке углеводородных цепей и полярных головок такие же, как и в липидных крастал-лах. Эти принципы будут обсуждаться в разд. 2.3 при анализе формы мицелл.
Многие методы, включая дифракцию рентгеновских лучей и нейтронов, спектроскопию КР и ИК-спектрометрию, указывают, что в фазе геля насыщенные углеводородные цепи фосфолипидов находятся преимущественно в полностью-гпрднс-конфигурации. Минимальная площадь поперечного сечения молекулы диа-цильного фосфолипида равна около 38 A2. Примерно такую площадь занимает полярная головка фосфатидилэтанола-мина, поэтому насыщенные фосфатидилэтаноламины в гелевой фазе упаковываются так, что ацильные цепи располагаются перпендикулярно плоскости бислоя, как и в липидных кристаллах. В случае же кристаллов фосфатидилхолина минимальная площадь, приходящаяся на одну полярную головку, составляет примерно 50 А2. Поэтому дипальмитоилфосфатидилхолин в гелевой фазе не может упаковываться так, как фосфатидилэтаноламин. В этом случае ацильные цепи дипальмитоилфосфатидилхолина отклоняются на 30° от нормали к бислою, благодаря чему их поперечное сечение увеличивается и достигается соответствие размеру полярной головки. При этом углеводородные цепи сохраняют полнос-тью-транс-конфигурацию. В жидкокристаллической фазе появление в цепи гош-конформеров увеличивает эффективное поперечное сече-

ние цепей по меньшей мере до 50 А2 в расчете на молекулу диациль-ного фосфолипида, а в водных дисперсиях эффективная площадь, приходящаяся на молекулу фосфолипида, составляет обычно 60 — 70 А2. Следовательно, в жидкокристаллической фазе углеводородные цепи не наклонены к плоскости бислоя, поскольку в этих условиях.полярные головки липидных молекул достаточно удалены друг от друга, и чтобы заполнить пространство между соседними головками и перекинуть между ними мостики, требуются вода и другие полярные молекулы. Судя по данным 2Н-ЯМР, толщина углеводородной области дипальмитоилфосфатидилхолинового бислоя в жидкокристаллическом состоянии составляет 35, а не 45 А, как следовало ожидать, если бы цепи находились в полностью-/я/*7«с-конфиругации и были ориентированы вдоль нормали к би-слою. Толщина бислоя уменьшается за счет наличия к цепях гош-конформеров, приводящих к разупорядоченности цепей, причем сами цепи в целом растянуты и расположены перпендикулярно поверхности бислоя, а не скручены в спираль. На рис. 2.12 приведена линейная зависимость толщины жидкокристаллического бислоя в диацилфосфатидилхолиновых везикулах от длины ацильных цепей, построенная по данным рассеяния рентгеновских лучей.
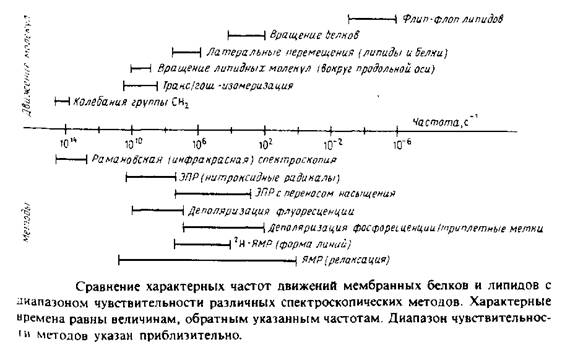
МЕТОДЫ ИССЛЕДОВАНИЯ ГИДРОФОБНОЙ ОБЛАСТИ БИСЛОЯ
Для получения детальной картины строения внутренней области бислоя особенно полезными оказались два метода: Н-ЯМР и ИК-и КР-спектроскопия. Применение *Н- 13С-ЯМР для исследования мембран и водно-липидных систем затруднено, поскольку для получения пригодных для исследования малых везикул или мембранных фрагментов необходима предварительная фрагментация мембранного препарата с помощью ультразвука. Впрочем, если вращать образец под магическим углом, то необходимость в такой обработке отпадает, и это открывает новые возможности для более широкого применения последних методологических достижений ЯМР при изучении модельных и биологических мембран.
Применение методов 2Н-ЯМР и колебательной спектроскопии не сопровождается возмущениями бислоя, поскольку в этих методах не используются зонды, которые могли бы исказить структуру окружающих их липидов. Ниже приведены краткое описание этих методов и сводка результатов, полученных при изучении конфигурации ацильных цепей.
Спектроскопия комбинационного рассеяния
Лазерная спектроскопия комбинационного рассеяния оказалась весьма полезной для изучения физического состояния модельных и биологических мембран. Этот метод основан на измерении разности энергии падающего света и света, рассеянного за счет колебательных движений. Тип колебаний и их интенсивность очень сильно зависят от физического состояния липидов, поэтому спектры фосфолипидов в фазе геля и жидкокристаллическом состоянии существенно различаются. Особенно полезны для регистрации этих различий валентные колебания связи С—С. Например, появление гош-конформеров при плавлении углеводородных цепей приводит к увеличению интенсивности полосы при 1080 см-1. Этот метод показывает, что некоторая доля гош-конформеров сохраняется и в состоянии геля до тех пор, пока температура не снизится до очень низких значений, около - 200° С. Заметим, что колебания часто охватывают всю молекулу, так что количественная интерпретация спектров с точки зрения анализа влияния отдельных коиформаций является непростой задачей.
Обычно образцы представляют собой суспензию липидов в концентрации около 1 мг/мл. Мембраны, содержащие хромофоры или флуоресцирующие примеси, непригодны для изучения. Фоновая флуоресценция делает невозможным измерение относительно слабых сигналов комбинационного рассеяния, а поглощение хромофорами лазерного излучения приводит к нагреванию
Инфракрасная спектроскопия
Этот метод также основан на регистрации колебательных спектров молекул, но до недавнего времени его применение для изучения биологических объектов было ограниченным из-за невозможности работы с водными суспензиями. После разработки современных ИК-спектрометров с фурье-преобразованием многие из этих проблем были решены. В настоящее время опубликовано немало работ по изучению липидных дисперсий и биологических мембран методом фурье-ИК-спектроскопии. Преимущества этого метода перед спектроскопией КР состоят в его значительно более высокой чувствительности, а также в том, что флуоресцирующие примеси или хромофоры не мешают измерениям. Как и в случае спектроскопии КР, фурье-ИК-спектры чувствительны к изменениям полиморф-ио-фазового состояния липидов. Поэтому фурье-ИК-спектроскопия использовалась для изучения предпереходов в фосфатидилхолино-вых бислоях, главного фазового перехода гель—жидкий кристалл, а также перехода яичного фосфатидилэтаноламина из ламеллярной фазы в гексагональную Нц-фазу.
Изменение коиформации липидных цепей сопровождается частотным сдвигом полос поглощения групп СНг, причем эти изменения коррелируют с изменением доли гош-коиформеров в цепях. Например, в присутствии холестерола число гош-коиформеров в ди-пальмитоилфосфатидилхолине при температурах выше температуры главного фазового перехода снижается, что согласуется с изменениями упорядоченности бислоя, измеренной методом 2Н-ЯМР. Встраивание в бислой интегрального мембранного белка оказывает совершенно иной эффект: число гош-кон-формеров в жидкокристаллической фазе практически не изменяется, но увеличивается их содержание в фазе геля, поскольку белки мешают ацильным цепям упаковываться в полностью-транс-конфигура-ции.
Более детальную картину строения гидрофобной области липидного бислоя удалось получить с помощью метода 2Н-ЯМР. Атомы водорода в определенных местах липидной молекулы можно избирательно заменить дейтерием. Это сравнительно мяг-
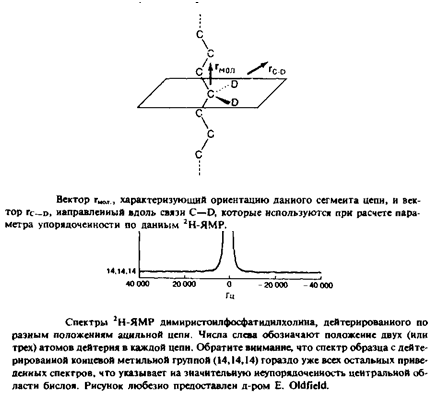
кий способ зондирования мембран, и считается, что он, как правило, не вносит возмущений в их структуру. Спектры некоторых дейтерированных димиристоилфосфатидилхолинов представлены на рис. Расстояние между двумя пиками Avq, называемое квадрупольным расщеплением, зависит от усредненной по времени ориентации вектора С—D-связи по отношению к нормали к бислою. Усредненную по времени ориентацию можно выразить через параметр упорядоченности следующим образом:
![]()
где <cos20) отражает усреднение ориентации по времени, a Scd является параметром упорядоченности связи. Необходимо подчеркнуть, что результат измерения является величиной, усредненной по всем молекулам.
![]()
При хаотичной ориентации. Этот параметр описывает усредненную ориентацию данного сегмента ацильной цепи:
мол = _ 2Scd-
Параметр упорядоченности, получаемый с помощью метода 2Н-ЯМР, отражает усредненную ориентацию и мало что говорит о динамике системы и о характере движений.
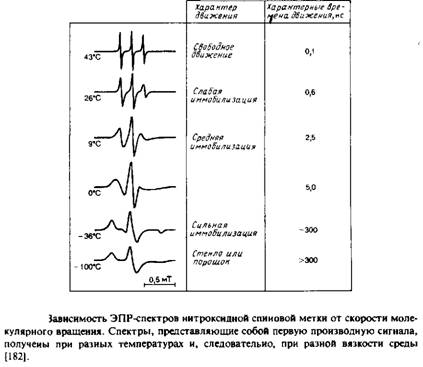
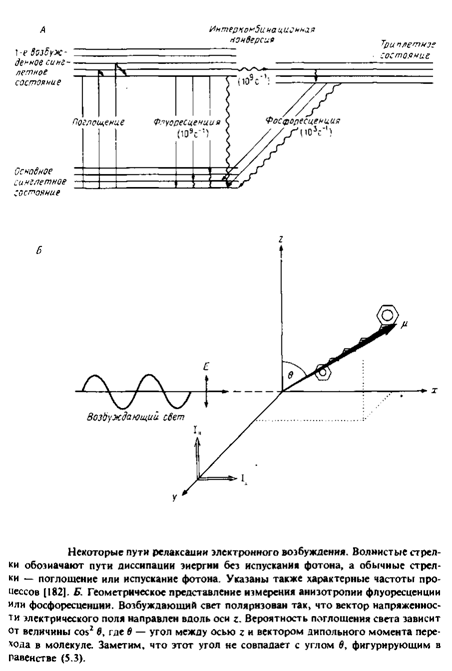
Особое значение имеет тот факт, что локальное магнитное поле, в котором находится конкретный атом дейтерия, зависит от ориентации С—D-связи по отношению к внешнему магнитному полю. Колебательные и вращательные движения молекулы, которые влияют на ориентацию С—D-связи в бислое в целом, происходят с достаточно большой скоростью, так что любой атом дейтерия воспринимает единое усредненное магнитное окружение. Это окружение зависит от соседних атомов, а также от ограничений движения по типу и амплитуде. В этом отношении метод 2Н-ЯМР отличается от КР- и ИК-спектроскопии, поскольку переходы транс-и гош- совершаются с гораздо меньшей частотой, чем разность частот колебательных полос, отвечающих этим формам. Поэтому ИК- и КР-спектры дают картину, которую можно назвать моментальной фотографией вклада транс- и гош-ротамеров в спектральные параметры. Для измерения параметров упорядоченности применяют и другие методы, в частности спектроскопию ЭПР и флуоресенцентную спектроскопию.
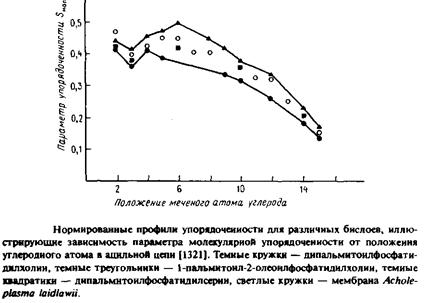
На рис. приведены параметры упорядоченности, определенные по данным 2Н-ЯМР для нескольких селективно дейтерирован-ных фосфолипидов, в которых атом дейтерия включен в определенные метиленовые группы sn-1-пальмитоильного остатка. Исследовались как липидные бислои, так и природные мембраны, находящиеся в жидкокристаллическом состоянии, поскольку в случае фазы геля спектры очень уширяются из-за плотной упаковки липидов и потому с трудом поддаются анализу. Эти данные позволяют сделать два вывода.
1.Параметр упорядоченности довольно постоянен на участке от С-2 и до примерно С-8 или С-10. Метиленовые группы в средней части бислоя значительно более разупорядочены, чем группы вблизи его поверхности.
2.Для синтетических липидов разных типов, включая фосфати-дилхолин, фосфатидилсерин и сфингомиелин, а также для биологических мембран, содержащих дейтерированные зонды, получен одинаковый профиль параметра упорядоченности. Таким образом, характер упорядоченности бислоя мало зависит от химического строения липида и от состава мембраны, если бислой находится в жидкокристаллическом состоянии.
Количественный анализ этих данных можно провести на основе молекулярного моделирования с использованием методов статистической механики. Например, приведенные результаты согласуются с наличием в каждой цепи дипаль-митоилфосфатидилхолина четырех или пяти гош-ротамеров при очень малом содержании кинков. Поскольку каждая цепь закреплена у поверхности бислоя, участок цепи вблизи поверхности наиболее упорядочен. Обратите внимание, что бислой — это высококооперативная система. Ацильная цепь не может изменить свое направление без компенсационных изменений соседних цепей. Поэтому группа смежных сегментов цепи должна двигаться кооперативно. Отклонения ориентации сегментов цепи от нормали к бислою будут усиливаться по мере перехода от поверхности бислоя к его центральной области. Поэтому разупорядочен-ность максимальна в середине бислоя, где подвижность цепей такая же, как в жидком парафине. Данные других методов также показывают, что молекулярная подвижность максимальна в центре бислоя. Но необходимо отметить, что неупорядоченность — это статистический параметр, который ничего не говорит о характере движения. Так, можно иметь сильно неупорядоченную структуру, которая в то же время обладает малой подвижностью.
Дифракция нейтронов
Селективно дейтерированные фосфолипиды можно также исследовать методом дифракции нейтронов. Если рентгеновские лучи рассеиваются на электронах, то нейтроны — на ядрах атомов. Рассеивающие свойства 'Н и 2Н сильно различаются, что позволяет выявлять дейтерированные области по кривой плотности рассеяния. Например, судя по данным дифракции нейтронов, атом С-5 находится на расстоянии 15 А от центра дипальмитоилфосфатидил-холинового бислоя в фазе геля; это соответствует полностью вытянутой транс-конфигурации цепей. Используя D2O в качестве растворителя, можно определить локализацию воды в фазе геля; полученные данные показывают, что вода проникает до области локализации глицерольных остатков. С помощью рассеяния нейтронов можно определить также локализацию ионов, таких, как Са2+.
Похожие работы
... является курсом, для изучения которого необходимо наличие знаний об основных принципах организации биологических молекул, строении и механизмах действия ферментов. Дисциплина биохимия мембран относится к дисциплинам специализации федерального компонента. 5. Распределение времени, отведенного на изучение дисциплины по учебному плану Форма учебной работы Форма обучения Очная По ...
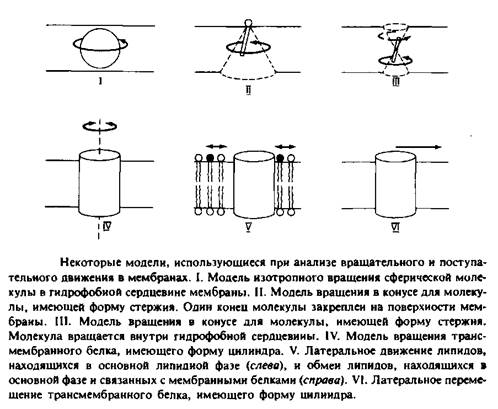
... к уменьшению латеральной диффузии. 5. Липидно-белковые взаимодействия Большинство методов, применяемых для изучения упорядоченности и динамических свойств мембран, используется и для исследования липидно-белковых взаимодействий. Работы по изучению этих взаимодействий были в основном направлены на выяснение влияния мембранных белков на физическое состояние липидов. Рассмотрим типичную мембрану ...



... животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы. Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. ...
... - восстановительные. 5.Ферменты патогенности (вирулентности). Ферментный состав клетки определяется геномом и является достаточно постоянным признаком. Знание биохимических свойств микроорганизмов позволяет идентифицировать их по набору ферментов. Основные продукты ферментирования углеводов и белков- кислота, газ, индол, сероводород, хотя реальный спектр для различных микроорганизмов намного ...
0 комментариев