Особенности роста микроскопических грибов в стандартных условиях культивирования
Особенности использования микромицетами различных природных веществ в качестве единственного источника углерода
Разложение трудноразлагаемых веществ
Объекты исследований
Определение способности плесневых грибов использовать различные источники углерода
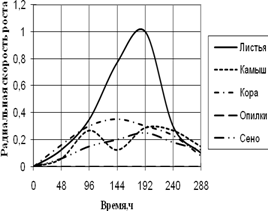
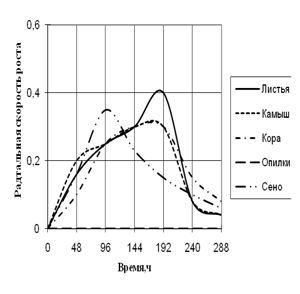
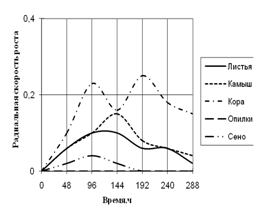
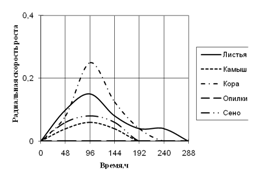
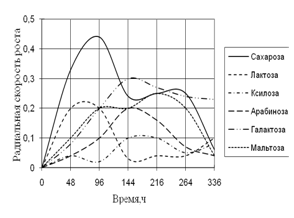
Радиальная скорость грибов на модельных субстратах
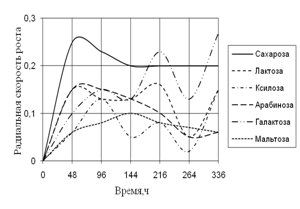
Радиальная скорость роста грибов на естественных субстратах
Прорастание спор в капле субстрата
Навигация
Исследование роста микромицетов на различных субстратах
Исследование роста микромицетов на различных субстратах
80698
знаков
6
таблиц
19
изображений
ВВЕДЕНИЕ
Микроскопические грибы значительно различаются способностью усваивать разные соединения углерода и синтезировать из них составные части клетки. Некоторые виды могут использовать для питания разнообразные соединения. С другой стороны, известно множество различных специализированных типов микромицетов, которые нуждаются в специфических соединениях. Нефть, газообразные углеводороды, парафин, воск, резины, гудрон, капрон и многие другие синтетические материалы, а также пестициды после попадания в почву начинают разлагаться плесневыми грибами и бактериями. Практически не существует органических соединений, которые не усваивались бы микроорганизмами.
Роль грибов в экологических системах велика и разнообразна. В трофических цепях они являются консументами, вторичными или третичными, но в основном выполняют функции редуцентов, минерализуют органические вещества растительного и животного происхождения. Мощные фотосинтетические системы в биосфере компенсированы столь же мощными и многообразными гидролитическими системами (ферментами) микробного мира. Микроскопические грибы являются важнейшими компонентами этой каталитической системы, способной трансформировать практически все природные органические соединения. Возможность использовать в качестве единственного источника углерода природные биополимеры определяется способностью грибов синтезировать соответствующие гидролитические экзоферменты, расщепляющие нерастворимые полимеры до мономеров, способных проникать в клетку.
Разнообразная группа органотрофных организмов, грибов и бактерий, осуществляющих деструкционную ветвь в цикле углерода, характеризуется кооперативными отношениями, при которых грибы-редуценты включаются в процесс после того, как легко разлагаемые органические вещества уже использованы бактериями (Домарадский, 2007).
Любое органическое вещество вначале расщепляется до более простых соединений, а последние вовлекаются в тот или иной биосинтетический процесс. Возникающая при расщеплении энергия накапливается в АТФ или в других соединениях, имеющих макроэргические связи. Таким образом, органические субстраты обеспечивают как энергетическую, так и конструктивную стороны обмена плесневых грибов и бактерий. Следовательно, вопрос об углеродном питании и о влиянии источников углерода на развитие микроорганизмов является очень существенным при проведении любых микробиологических экспериментов (Лилли, 1957).
Целью данной работы является изучение особенностей использования чистыми культурами микромицетов различных источников углерода.
В соответствии с поставленной целью задачами исследования является определение у коллекционных штаммов микроскопических грибов:
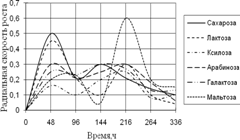
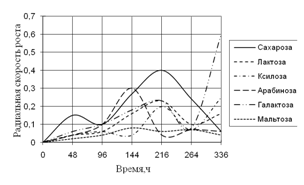
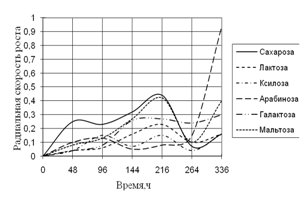
1) способности использовать модельные источники углерода (сахара, многоатомные спирты, крахмал, целлюлозу);
2) способности к росту на природных субстратах (растительный опад, камыш, сено, опилки, кора) по радиальной скорости роста;
3) способности к ассимиляции природных субстратов (растительный опад, камыш, сено, опилки, кора) на основе метода проращивания грибных зачатков во влажной камере.
Практическая значимость работы заключается в том, что в работе изучены биохимические особенности 10 штаммов коллекционных микромицетов. Полученные данные могут служить основой составления паспортов штаммов.
ГЛАВА 1. ОСОБЕННОСТИ МИКРОСКОПИЧЕСКИХ ГРИБОВ, ВЫДЕЛЕННЫХ ИЗ РАЗЛИЧНЫХ ЭКОНИШ
1.1 Влияние экологических условий на рост микромицетов
1.1.1 Сравнительный анализ скорости радиального роста микромицетов, выделенных из различных экотопов
На сегодня экологические проблемы становятся не менее актуальными, чем запросы микробиологической промышленности (Кочкина, 1978; Великанов, 1983; Жданова, 1982-2000; Паников, 1991). Надежды на их решение связывают с так называемым системным подходом, который предполагает использование количественной информации о природных объектах (Паников, 1991). Здесь чрезвычайно остро ощущается недостаток знаний в отношении почвенных микроорганизмов (Гильманов, 1983; Parton, 1988), и большая часть дефицита информации относится именно к вопросам роста микробных популяций (Паников, 1991). Этот недостаток обусловлен тем, что фундаментальные основы и принципы микробиологических исследований вырабатывались в отрыве от решения экологических задач. По данным ряда авторов (Кочкина, 1978; Паников, 1991), экологически наиболее корректно выращивание грибных колоний на агаризованных средах, так как пространственно-неоднородный рост микроскопических грибов наиболее приближен к их росту в природных условиях.
Таким образом, изучение ростовых процессов способствует описанию многообразия адаптивных поведенческих реакций у микроорганизмов в изменяющихся условиях среды (что важно для экологии) (Паников, 1991).
Известно, что под действием неблагоприятных факторов, в том числе и радиационного загрязнения, у организмов происходят глубокие изменения, которые проявляются на морфологическом, физиологическом и биохимическом уровнях. В то же время такой физиологический показатель, как скорость роста, является достаточно консервативным для каждого вида микромицетов (Кочкина, 1978).
В работах последних лет было показано, что при длительном существовании микромицетов на высокорадиоактивных субстратах радиальная скорость роста некоторых видов существенно изменяется (Жданова, 2000; Вембер, 2001). Для того чтобы проанализировать, у каких именно видов происходят подобные изменения (в зависимости от их систематического положения и морфологических особенностей), Блажеевской с соавторами проведено сравнительное изучение радиальной скорости роста у различных видов микромицетов. Часть штаммов каждого из видов была представлена грибами, выделенными из радиоактивно загрязненных местообитаний, а другая часть – из радиоактивно чистых экотопов (Блажеевская, 2002).
Скорости роста изученных штаммов грибов оценивали с учетом места их выделения и характера среды культивирования.
По скорости радиального роста все изученные виды можно условно разделить на три основные группы: 1) виды с наивысшей скоростью роста – Ulocladium consortiale и Alternaria alternatа; 2) вид Cladosporium sphaerospermum, скорость роста которого была наиболее низкой; 3) остальные изученные виды занимали промежуточное положение по этому параметру (Блажеевская, 2002).
Большое значение заслуживает характер ритмических колебаний роста изученных культур. Биологические ритмы присущи всем уровням живой материи – от молекулярных и субклеточных структур до биосферы, что обеспечивает единство живой и неживой природы. Все они отражают процессы регулирования функций организма (Романов, 1980).
Биохимический механизм циркадных ритмов связывают с обменом нуклеиновых кислот (Шаркова, 1971). Имеющиеся данные указывают на то, что с усилением ритма инициируется обмен РНК. Биоритмы митоспоровых грибов отражают неограниченно восстанавливаемый адаптивный онтогенез, формирующийся на основе сигналов окружающей среды, генетический механизм которого базируется на высокой способности грибов к формированию многоядерности и ядерного дуализма (Беккер, 1983).
Характер и продолжительность ростовых ритмов, так же как и радиальная скорость роста, зависят от систематического положения гриба, его возраста (возраста культуры), концентрации в среде основных источников питания и энергии, а также других экологических факторов (Романов, 1980; Бухало, 1988). Ритмы грибов варьируют от 4 – 5 часов до 7 – 9 суток, нескольких недель и даже лет (Шаркова, 1971).
Блажеевской Ю. В. было показано, что штаммы с замедленной ритмичностью (>1-суточных) выделены из наиболее радиоактивно загрязненных местообитаний. Кроме того, среди штаммов с наибольшей продолжительностью биоритмов (1 – 2-суточные) преобладали штаммы с невысокой скоростью роста. В подавляющем большинстве случаев изученные виды примерно одинаково росли на голодной среде и сусло-агаре. В чистых и загрязненных почвах нет заметной разницы между количеством видов, предпочитающих богатую среду, и количеством видов, одинаково растущих на обеих средах. Замедление ростовых процессов отмечено только у штаммов, долгое время существовавших на субстратах с высоким уровнем радиоактивного загрязнения. Подобное замедление скорости роста может свидетельствовать об адаптивной перестройке метаболических процессов у штаммов, длительное время растущих на высокорадиоактивных субстратах (Блажеевская, 2002).
Похожие работы
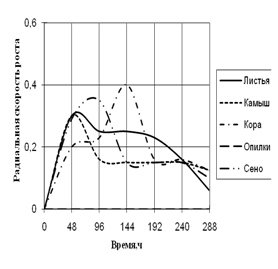
... штаммов использовать в качестве единственного источника углерода природные растительные материалы (камыш, сено, растительный опад, опилки, кора). 3. Исследование роста микромицетов на различных источниках углеродного питания Объектами исследования явились коллекционные штаммы микроскоскопических родов Aspergillus flavus, A. fumigatus, Alternaria sp., Penicillium sp., Cladosporium ...
... 5 видов разлагали хитин, 8 – выделяли тирозиназу и лакказу (Parton, 1988). Отмечают высокую целюлозолитическую активность у штаммов видов Trichoderma, Aspergillus niger и Penicillium miszynsky, выделенных из водных местообитаний (Hopper, 1995). Микроскопические грибы могут прямо или косвенно влиять на активность и развитие других гидробионтов. Пути воздействия могут быть различными – от прямого ...

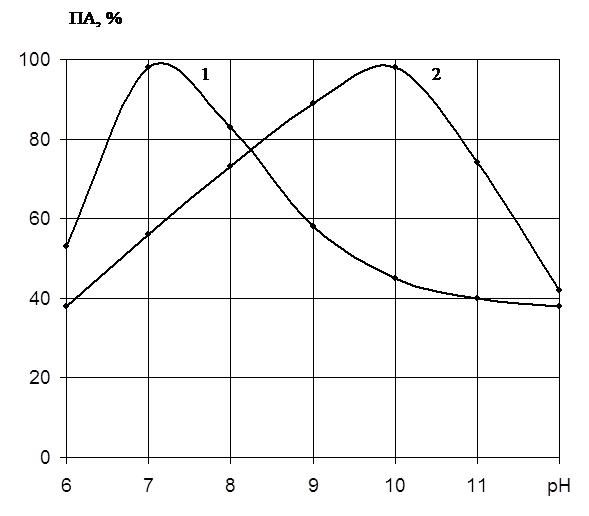
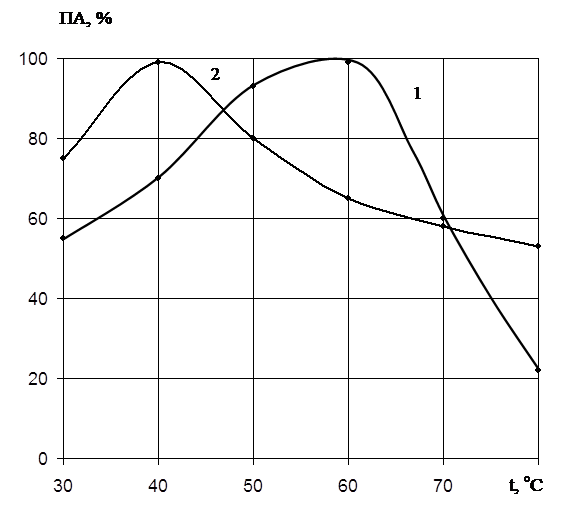
... есть среднее из двух или трех определений. Обсуждаются только те результаты, которые были воспроизводимы в каждом опыте. ГЛАВА 3. ПОЛУЧЕНИЕ ПРЕПАРАТОВ ПРОТЕИНАЗЫ PENICILLIUM WORTMANNII 2091 И ИССЛЕДОВАНИЕ ИХ ФИЗИКО-ХИМИЧЕСКИХ СВОЙСТВ. Известно, что микроорганизмы синтезируют богатые набором ферментов комплексы. Поэтому важным этапом в получении препаратов направленного действия является ...
... исследуется лишь суммарное содержание веществ либо число компонентов, не превышающее 3-5). Данные по процессам, идущим при биодеградации и фотолизе многокомпонентных (более 10-ти веществ) смесей полиароматических углеводородов отсутствуют как в отечественной так и зарубежной литературе. Резюмируя вышеперечисленное можно сказать, что сегодня в России и развитых зарубежных странах ведутся ...
0 комментариев