Особенности роста микроскопических грибов в стандартных условиях культивирования
Особенности использования микромицетами различных природных веществ в качестве единственного источника углерода
Разложение трудноразлагаемых веществ
Объекты исследований
Определение способности плесневых грибов использовать различные источники углерода
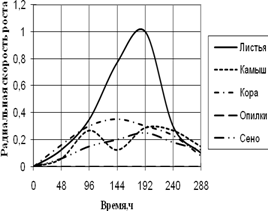
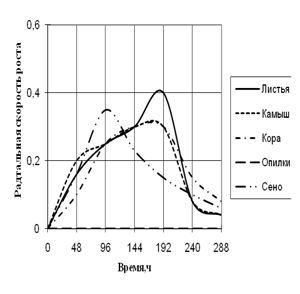
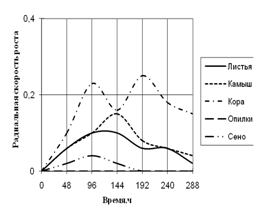
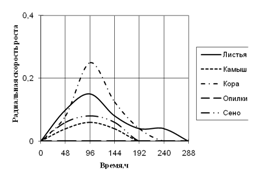
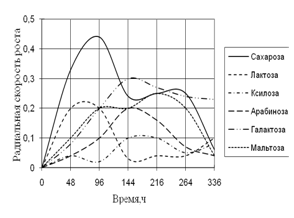
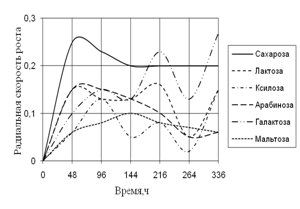
Радиальная скорость грибов на модельных субстратах
Радиальная скорость роста грибов на естественных субстратах
Прорастание спор в капле субстрата
Навигация
Разложение трудноразлагаемых веществ
Исследование роста микромицетов на различных субстратах
80698
знаков
6
таблиц
19
изображений
1.2.2 Разложение трудноразлагаемых веществ
Плесневые грибы в качестве источника углерода могут использовать такие трудноразлагаемые вещества, как целлюлоза, крахмал, лигнин, пектиновые вещества, нефть, пестициды.
Разложение целлюлозы. Основными источниками целлюлозы для грибов в природных условиях служат древесина и различные растительные остатки. Среди микроскопических грибов, разлагающих целлюлозу известны представители следующих родов: Aspergillus, Coriolus, Eupenicillium, Fusarium, Penicillium, Sporotrichum, Trichoderma, Verticillium. Но только некоторые из них продуцируют полные внеклеточные целлюлолитические системы (эндо- и экзоглюканазы β-глюкозидазу). Среди них Trichoderma viride, T. reesei, T. koningii, Penicillium funiculosum, Fusarium solani. Для культуральной жидкости большинства других грибов этой группы характерно отсутствие экзоглюканазы, то есть эти грибы могут деградировать более аморфные формы целлюлозы.
Деградация высокоупорядоченной формы целлюлозы осуществляется благодаря синергическому действию комплекса целлюлолитических ферментов. При любой комбинации экзо- и эндоглюканаз Trichoderma koningii, Fusarium solani, Penicillium и Funiculosum отмечается выраженный синергизм. Однако синергизм между экзоглюканазами этих грибов и эндоглюканазами грибов, не продуцирующих экзоглюканазу (Myrothecium verrucaria), не выявлен. Нет также синергизма между экзоглюканазами грибов и эндоглюканазами рубцовых бактерий. Последнее указывает на существенные различия целлюлолитических систем грибов и бактерий (Марьиновская, 2006).
Целлюлоза является линейным полимером d-глюкозы. Остатки глюкозы в молекуле клетчатки, как и в молекуле целлобиозы связаны β-гликозидной связью. Поэтому клетчатку можно рассматривать как полимер целлобиозы. Нормэн и Фуллер (1942) считают, что большинство грибов способно усваивать клетчатку. Несмотря на то, что использование клетчатки грибами имеет большое значение в круговороте веществ в природе, процесс этот изучен далеко не полно.
Обычно считают, что первым этапом использования целлюлозы грибами является ее гидролиз. Гидролиз целлюлозы можно схематически представить следующим образом: целлюлоза → целлодекстрины → целлотетроза → целлобиоза → d-глюкоза. Известен штамм Aspergillus oryzae, выделяющий целлюлазу и целлобиозу (Лилли, 1957).
Разложение крахмала. Как и целлюлоза, крахмал является полимером d-глюкозы. Остатки глюкозы в его молекуле соединены между собой α-гликозидной связью, поэтому основной структурной единицей молекулы крахмала, как и молекулы гликогена, следует считать мальтозу. Зеленые растения синтезируют крахмал, животные и грибы образуют гликоген. Ферментативный гидролиз крахмала может быть схематически представлен следующим образом: крахмал→декстрины→мальтоза→d-глюкоза. Декстрины, имеющие разветвленную углеродную цепочку, лишь частично гидролизуются амилазой. Декстрины с неразветвленной углеродной цепочкой полностью превращаются в мальтозу (Мирчинк, 1988).
Крахмал нерастворим в воде. Лишь грибы, образующие амилазу, обладают способностью усваивать крахмал. Существует немало грибов, неспособных развиваться на средах с крахмалом, однако большинство из них может усваивать этот полисахарид. Было установлено, что 26 различных изученных видов и штаммов оомицетов из числа сапролегниевых усваивали как крахмал, так и продукты его гидролиза (декстрины, мальтозу и глюкозу), но не были способны ассимилировать 13 других источников углерода, включая сюда и фруктозу. Позднее А. С. Марголин (1942) показал, что 19 из 21 вида грибов, усваивающих мальтозу, обладали также способностью использовать и декстрин (Лилли, 1957).
Разложение пектиновых веществ. Среди грибов имеются активные разлагатели пектина, который является существенным компонентом растительного опада. Пектин образует в растениях межклеточное вещество, из которого состоят так называемые срединные пластинки, соединяющие между собой отдельные клетки растения. Они придают тканям прочность. Пектин представляет собой высокомолекулярное соединение углеводной природы – полисахарид, в котором метоксилированные остатки галактуроновой кислоты связаны между собой β-1,4-глюкозидными связями.
Многие грибы образуют пектинолитические ферменты. Высокая пектинолитическая активность обнаружена у некоторых эпифитных грибов, главным образом Aureobasidium pullulans и видов Cladosporium. Пектинолитические грибы занимают значительное место среди типичных представителей лесной подстилки – это виды родов Cladosporium, Alternaria, Aposphaeria, Penicillium, фитопатогенные грибы родов Fusarium, Verticillium, Botrytis cinerea, Sclerotinia sclerotiorum.
Ферментативное разрушение пектиновых веществ в растениях имеет значение в патогенезе некоторых заболеваний. Фитопатогенные грибы разрушают пектин срединной пластинкой и пектаты в первичных клеточных оболочках, что приводит к изменению их физико-химических свойств и создает условия для внедрения паразита, а также в результате действия пектинэстеразы образуются вещества – полигалактурониды, способные закупоривать сосуды, что в конечном итоге приводит к увяданию растений.
Существенное значение разрушения пектиновых веществ грибами имеет при разложении растительного опада. Практическое использование пектиназ грибов – применение в пищевой промышленности при приготовлении фруктовых соков для их осветления, а также при мочке льна (Мирчинк, 1988).
Разложение лигнина. Грибы – почти единственные разрушители лигнина. Способность грибов осуществлять глубокое разрушение лигнина представляет собой уникальное явление.
Лигнин – наиболее распространенное в природе полимерное циклическое соединение. В наибольшем количестве лигнин содержится в древесине и древесном опаде. Содержание его в опаде хвойных пород составляет 28 – 34 %, лиственный пород – 18 – 28 %. В химическом отношении лигнин не является индивидуальным веществом с вполне определенными свойствами и составом.
Исследования, касающиеся микробной деградации лигнина, относятся к одной из наиболее сложных биологических проблем, поскольку лигнин пока не может быть точно определен как химическое вещество. Также не могут быть точно определены и промежуточные реакции его биологического превращения. Наиболее активные группы микроорганизмов, разрушающих лигнин, принадлежат к древоразрушающим базидиомицетам, вызывающим белую гниль. Однако до настоящего времени неизвестны полностью все стадии ферментативных реакций в процессе разложения лигнина, то есть известны далеко не все ферменты, осуществляющие этот процесс.
Отличия деградации лигнина от деградации других полимеров заключается в том, что такие полимеры, как протеины, полисахариды, нуклеиновые кислоты, состоят из регулярно повторяющихся единиц, в то время как лигнин состоит из различных мономеров, имеющих различные типы связей. Большинство микроорганизмов, воздействующих на лигнин, вызывают в нем очень незначительные изменения, которые проявляются в основном в уменьшении числа метоксильных групп и очень слабой потере в весе. Некоторые сумчатые и несовершенные грибы могут расти на средах, содержащих препарат лигнина в качестве единственного источника углерода, такие, как Fusarium lactis, F. nivale и некоторые другие, но они не вызывают существенных изменений в молекуле лигнина (Мирчинк, 1988):
Участвуя в разложении многих углеродсодержащих веществ растительного опада и древесины в первую очередь трудноразлагаемых полимерных соединений, где грибам принадлежит ведущая роль, они занимают значительное место в круговороте углерода, являясь поставщиками СО2 в атмосферу.
Среди грибов есть организмы, разлагающие жиры и воска, входящие в состав растительных и животных тканей. Это определяется наличием у них ферментов липаз. Наибольшей активностью липолитических ферментов обладают виды Mucor lipolyticus, Rhizopus nigricans, Aspergillus niger, Penicillium verrusum, Penicillium roquefortii. Многие выделены с поверхности растений, являясь эпифитами и способны разлагать также восковые налеты на поверхности растений.
Известна также способность грибов разлагать как алифатические, так и ароматические углеводороды. В этом отношении наибольшей активностью характеризуются грибы рода Aspergillus (Мирчинк, 1988).
Разложение хитина. Хитин постоянно присутствует в почве, достигая десятых долей процентов, входит в состав наружного скелета беспозвоночных животных и клеточных стенок грибов. По ряду физико-химических свойств хитин сходен с целлюлозой, однако наличие в молекуле ацетамидных групп придает ему особо ценные в практическом отношении свойства. Известно, например, что бактериальная хитиназа используется в качестве средства защиты растений от возбудителей болезней. Препарат на основе этого фермента является перспективным экологически безопасным средством биологического контроля за фитопатогенными грибами (Deboer, 1998; Eltarabily, 2000). Микроорганизмы воздействуют на хитин с помощью экзоферментов (хитиназы и хитобиазы), в результате чего образуются хитотриозы и хитобиозы, расщепляющиеся затем до мономеров и N-ацетилглюкозоамина (Шлегель, 1987). Способностью к образованию хитиназы обладают многие бактерии (Актуганов, 2003; Ramirez, 2004). В первую очередь реагируют на присутствие хитина быстрым размножением мицелиальные прокариоты – актиномицеты (Калакуцкий, 1977; Schrempf, 2001). Наличие высокой хитиназной активности микроорганизмов дает возможность извлечения азота и углерода из труднодоступных соединений, каковым является хитин, и, как следствие, включение этих элементов в круговорот почва – атмосфера. Однако проблема разложения хитина в почве до настоящего времени остается недостаточно раскрытой (Манучарова, 2005).
Манучаровой Н. А. с соавторами было проведено исследование хитинолитического прокариотного и эукариотного микробного комплекса в черноземе в ходе сукцессии, инициированной внесением хитина и увлажнением почвы.
Наблюдение за динамикой хитинолитических популяций в ходе сукцессии, инициированной увлажнением почвы и внесением хитина, с помощью люминесцентной микроскопии показало, что численность бактерий, длина мицелия актиномицетов и грибов в варианте с внесением хитина была выше по сравнению с контрольным вариантом (увлажнением почвы без внесения хитина), причем такая закономерность наблюдалась на всех этапах сукцессии. Максимальные значения численности бактерий, длины мицелия грибов и актиномицетов отмечены на 7 – 14-е сутки после начала опыта. На протяжении всего эксперимента биомасса грибов превышала биомассу прокариот как в контрольном варианте, так и в варианте с хитином. Значительное возрастание биомассы эукариот в варианте с хитином по сравнению с контролем отмечалось на 7-е сутки сукцессии, однако к середине сукцессии (14-е сутки опыта) биомасса грибов начала снижаться, что, вероятно, связано с сопряженными процессами отмирания части мицелия и перехода к стадии спороношения.
Наблюдение за динамикой популяций в ходе сукцессии, инициированной увлажнением и внесением хитина в почву, проводимое методом посева, показало, что численность хитинолитических прокариотических микроорганизмов, выделяемых из образцов с добавлением хитина, была выше на всех этапах сукцессии по сравнению с контролем (Манучарова, 2005).
Разрушение грибами нефтепродуктов. В последние десятилетия в связи с возродившимся интересом к процессам микробного превращения углеводородов были обнаружены мицелиальные грибы, деятельность которых приводит к деструкции нефти и ее производных. В настоящее время доказано, что утилизировать нефтепродукты, в том числе различные топлива, во время хранения и транспортировки способны многие виды грибов и бактерий (Андреюк, 1980).
Нефтепродукты как среда обитания грибов характеризуются рядом особенностей: 1) содержат большое количество сравнительно доступного углерода и минимальное – азота при почти недоступном пространственном расположении его в молекуле; 2) в них почти отсутствует доступная активная вода. Это оказывает существенное влияние на синтез de novo грибной клетки.
Вопросы необходимого соотношения C:N у грибов при росте на нефтепродуктах в биохимическом аспекте исследованы еще мало и уровень этих данных уже не отвечает современным представлениям о возможностях грибной клетки. Очевидно, здесь имеет место не только типичный гетеротрофный процесс, но также определенное подобие хемотрофии и автотрофии, причем стадии роста отличаются и специфичны по способности к разным типам трофики. Особенно это проявляется в период формирования репродуктивных структур (Ниязова, 1982; Бабьева, 1983). Спецификой роста грибов на нефтепродуктах является их способность распространяться на поверхности, то есть возможность использовать при этом активную воду из воздуха, а также расти в толще нефтепродуктов, то есть ограничивать свои потребности в воде за счет активной воды самих нефтепродуктов (Евдокимова 1982).
Рост грибов (кладоспориев, пенициллиев, аспергиллов и некоторых других видов и штаммов) в разных нефтепродуктах характеризуется различным типом размещения мицелиальной пленки. Наиболее типичный – на разделе фаз, однако чаще всего наблюдается еще и глубинный рост, при котором развивается не только в толще жидкости – до 20 см. Причем интересно, что рост этих штаммов при определенном соотношении нефтепродуктов и воды мало зависит от высоты слоев смеси, а также воздуха в надсубстратном пространстве. Это свидетельствует о большой возможности мицелиальных грибов выдерживать жесткие условия и приспосабливаться к потреблению необходимых для метаболизма веществ не совсем обычными биохимическими и физиологическими путями.
В настоящее время установлено, что способность окислять углеводороды нефти не является специфической чертой отдельных видов грибов. Это не редкая их особенность, а одна из физиологических функций. Однако, несмотря на большое сходство химических и физических свойств фракций нефтепродуктов, у большинства видов грибов четко проявляется избирательное отношение к их утилизации (Бабьева, 1983).
Разрушение полимерных материалов. Синтез полимеров и создание на их основе материалов, обладающих повышенной стойкостью к факторам окружающей среды и воздействию различных организмов, привел к обострению экологической обстановки из-за накопления больших объемов отходов, содержащих эти соединения в разных отраслях промышленности. В последние десятилетия во многих странах уделяется большое внимание созданию полимерных материалов и их модификаций, утилизация которых возможна под воздействием микробиоты. В качестве добавок к пластификаторам исследователи используют природные компоненты такие, как крахмал, производные целлюлозы, протеин, хитозан и так далее. На основе этих композитных полимеров ряд фирм выпускает пластики для производства изделий разового пользования, упаковки пищевых продуктов, плоских пленок и так далее, которые обладают способностью к биодеградации при компостировании и так далее (Власова, 2001; Фомин, 2001). Состав микроорганизмов, контаминирующих техногенные материалы и способных вызывать их биодеградацию, очень разнообразен как в таксономическом отношении, так и по их физиолого-биохимической активности. Среди них ведущее место занимают представители дейтеромицетов, способные развиваться на обширном сортименте материалов, содержащих соединения как природного происхождения, так и искусственного синтеза (Биоповреждения, 1987; Коваль, 1989).
Было проведено исследование О. В. Сычуговой с соавторами (2003) с целью изучения возможности роста и развития видов микромицетов на композиции пленочного сополимера этилена и винилацетата с термопластичным крахмалом.
В процессе данного исследования было установлено, что они способны утилизировать данный источник углеродного питания. Однако динамика роста видов на разных средах при одинаковых условиях инкубации, при одной и той же навеске крахмала не одинакова, что особенно четко проявляется на 4 – 10-е сутки. Выявляется и некоторая разница в темпе роста видов грибов на нативном и растворимом крахмале разного происхождения, а также на средах Чапека и Гетченсона, взятых в качестве контроля (Сычугова, 2003).
Изменения морфологических признаков и образования новых структур у тест – культур на модифицированных средах при замене сахарозы на крахмал и росте на полимере не отмечаются, и они сопоставимы с параметрами, приведенными в определителях (Пидопличко, 1953; Ellis, 1971; Watanabe, 2000). Отличия выявлены у них только в темпах формирования морфологических структур. Хотя в литературе и приведены данные о влиянии субстрата на появление новых морфологических структур у грибов (Богомолова, 2001), однако, вероятнее всего, онтогенез и темпы развития определяются геномом вида, реализация программы которого зависит от влияния различных факторов (Шевцова, 1987; Долгова, 1997).
На поверхности пленки, содержащей термопластичный крахмал фиксируются сформированные пучки конидиеносцев Aspergillus niger, Paecilomyces variotii, Penicillium funiculosum, Chaetomium globosum, Trichoderma viride. Другие виды из взятого набора тест-культур не растут на данном субстрате или формируют слабое спороношение и в более поздние сроки.
Рост мицелия и формирование спороношения на композиции пленочного сополимера дает основание предполагать участие некоторых видов грибов в биодеструкции полимера из сополимеров этилена и винилацетата (СЭВА) с добавлением термопластичного крахмала (ТПК). Это дает возможность при последующих пересевах на смесях СЭВА и ТПК отобрать наиболее активные виды и их штаммы для разработки биотехнологии по его утилизации (Сычугова, 2003).
Таким образом, микроскопические грибы могут использовать в качестве источников углерода разнообразные органические вещества, тем самым являясь важными деструкторами различных природных материалов: целлюлозы, крахмала, лигнина, гемицеллюлозы, жиров, углеводородов, а также синтетических материалов, таких как пластики, пленки, упаковки пищевых продуктов. Поэтому предполагаемое исследование актуально и своевременно.
ГЛАВА 2. ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Похожие работы
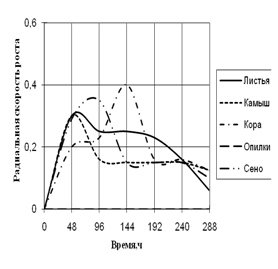
... штаммов использовать в качестве единственного источника углерода природные растительные материалы (камыш, сено, растительный опад, опилки, кора). 3. Исследование роста микромицетов на различных источниках углеродного питания Объектами исследования явились коллекционные штаммы микроскоскопических родов Aspergillus flavus, A. fumigatus, Alternaria sp., Penicillium sp., Cladosporium ...
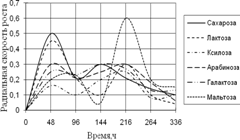
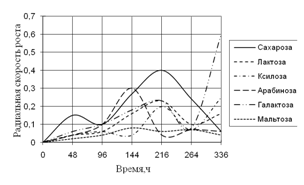
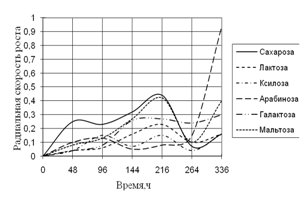
... 5 видов разлагали хитин, 8 – выделяли тирозиназу и лакказу (Parton, 1988). Отмечают высокую целюлозолитическую активность у штаммов видов Trichoderma, Aspergillus niger и Penicillium miszynsky, выделенных из водных местообитаний (Hopper, 1995). Микроскопические грибы могут прямо или косвенно влиять на активность и развитие других гидробионтов. Пути воздействия могут быть различными – от прямого ...

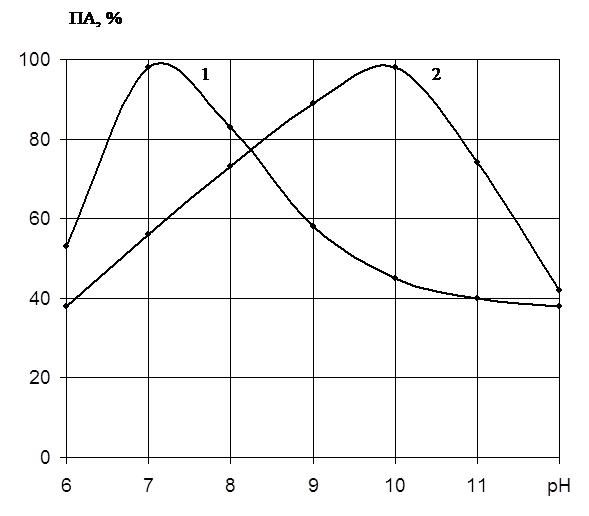
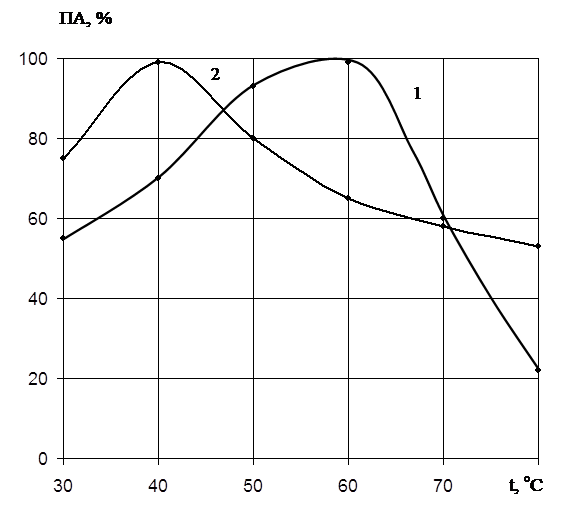
... есть среднее из двух или трех определений. Обсуждаются только те результаты, которые были воспроизводимы в каждом опыте. ГЛАВА 3. ПОЛУЧЕНИЕ ПРЕПАРАТОВ ПРОТЕИНАЗЫ PENICILLIUM WORTMANNII 2091 И ИССЛЕДОВАНИЕ ИХ ФИЗИКО-ХИМИЧЕСКИХ СВОЙСТВ. Известно, что микроорганизмы синтезируют богатые набором ферментов комплексы. Поэтому важным этапом в получении препаратов направленного действия является ...
... исследуется лишь суммарное содержание веществ либо число компонентов, не превышающее 3-5). Данные по процессам, идущим при биодеградации и фотолизе многокомпонентных (более 10-ти веществ) смесей полиароматических углеводородов отсутствуют как в отечественной так и зарубежной литературе. Резюмируя вышеперечисленное можно сказать, что сегодня в России и развитых зарубежных странах ведутся ...
0 комментариев