Навигация
Исследование роста микромицетов на различных источниках углеродного питания
56057
знаков
0
таблиц
7
изображений
Введение
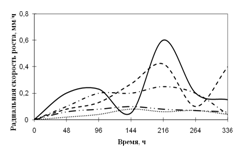
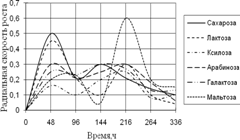
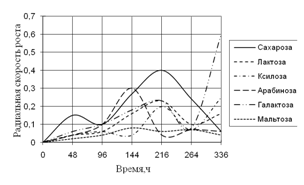
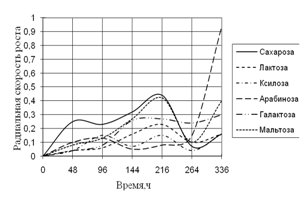
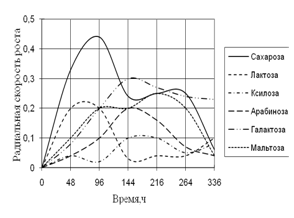
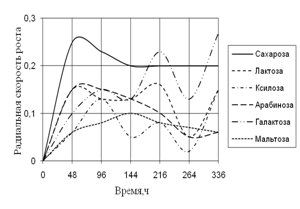
Среди необходимых для жизни микроскопических грибов элементов углерод занимает совершенно особое положение. Около половины сухого веса микромицета приходится на долю углерода. Этот элемент является постоянной составной частью протоплазмы, ферментов, клеточной оболочки и запасных питательных веществ. Поэтому соединения, содержащие углерод, играют важнейшую роль в питании микроскопических грибов (Лилли, 1957).
Микроскопические грибы значительно различаются способностью усваивать разные соединения углерода и синтезировать из них составные части клетки. Некоторые виды могут использовать для питания разнообразные соединения. С другой стороны, известно множество различных специализированных типов микромицетов, которые нуждаются в специфических соединениях. Нефть, газообразные углеводороды, парафин, воск, резины, гудрон, капрон и многие другие синтетические материалы, а также пестициды после попадания в почву начинают разлагаться плесневыми грибами и бактериями. Практически не существует органических соединений, которые не усваивались бы микроорганизмами.
Любое органическое вещество вначале расщепляется до более простых соединений, а последние вовлекаются в тот или иной биосинтетический процесс. Возникающая при расщеплении энергия накапливается в АТФ или в других соединениях, имеющих макроэргические связи. Таким образом, органические субстраты обеспечивают как энергетическую, так и конструктивную стороны обмена плесневых грибов и бактерий. Следовательно, вопрос об углеродном питании и о влиянии источников углерода на развитие микроорганизмов является очень существенным при проведении любых микробиологических экспериментов (Лилли, 1957).
Целью данной работы является изучение способности использования чистыми культурами микроскопических грибов природных растительных материалов в качестве источника углерода и энергии.
В соответствии с поставленной целью задачей исследования является:
1) определение радиальной скорости роста коллекционных микромицетов при росте на природных растительных материалов (растительный опад, камыш, сено, опилки, кора).
Практическая значимость работы заключается в том, что в настоящее время большой практический интерес вызывают микроорганизмы, в частности микроскопические грибы, участвующие в процессе деструкции природных растительных материалов.
1. Особенности использования углерода микромицетами
1.1 Использование углерода микроскопическими грибами
Соединения углерода используются грибами или как источники энергии, или же для синтеза важнейших составных частей клетки. Иногда на определенном этапе химических превращений оба эти процесса совпадают. Однако после образования некоторых промежуточных соединений, они обычно принимают различное направление. Определение общего количества усвоенного углерода не представляет больших затруднений. Значительные трудности встречаются, однако, при попытках проследить все химические превращения усвоенного соединения углерода в организме.
Баланс углерода. Общее представление о путях усвоения различных источников углерода может быть получено при определении количества выросшего мицелия, выделившейся углекислоты и других продуктов обмена веществ. Если первоначальное количество углерода известно, то его распределение в продуктах обмена веществ можно установить с помощью химических анализов. Обычно в этих опытах удается обнаружить 95 – 99% первоначального количества углерода.
Коэффициент использования. Связь между количеством продуктов обмена веществ, образованных грибов, и количеством использованного углеродистого или какого-либо другого соединения часто выражают математически, в виде отношения. Совершенно очевидно, что такого рода показатели (коэффициенты использования) представляют ценность лишь для данного вида гриба в определенных экспериментальных условиях. Иными словами, эти коэффициенты имеют абсолютное значение лишь для тех условий, в которых они были получены. Определение различных коэффициентов использования представляет не столь большую ценность, как установление полного баланса углерода, но зато оно требует значительно меньше химических анализов. Так как значение этих коэффициентов меняется в процессе развития гриба, то для получения более точных данных они должны определяться несколько раз, через различные интервалы времени после засева среды.
Наиболее широкое распространение из всех этих коэффициентов получил так называемый экономический коэффициент, выражающий соотношение между весом мицелия и спор и весом использованного сахара (или других источников углерода). Для этого по окончании опыта определяют оставшееся неусвоенным количество углерода в среде. Грибы, обладающие достаточной биохимической активностью, превращают приблизительно половину сахара, содержащегося в среде, в составные части клетки. Эффективность превращения веществ большинством грибов, культивируемых на лабораторных средах, значительно меньше. Это отчасти зависит от неправильного соотношения различных питательных веществ в среде и от характера углеродного обмена. Углерод, не использованный для синтеза мицелия, превращается в углекислоту и в промежуточные продукты обмена веществ, например спирт или органические кислоты. В промышленности стараются создать такие условия культивирования, при которых возможно большее количество углерода идет на образование нужного промежуточного продукта обмена и возможно меньшее количество расходуется на синтез мицелия и образование углекислоты.
Промежуточный обмен веществ. Усваиваемость различных источников углерода данным видом гриба определяется в основном структурой молекул этих соединений. Использование сложных природных соединений, например, полисахаридов, возможно лишь в том случае, если гриб способен синтезировать и выделять в среду необходимые гидролитические ферменты. Использование простых соединений, например, моносахаридов, также состоит из ряда следующих одна за другой ферментативных реакций обмена веществ. Известно, что химический состав гриба почти не зависит от использованного им источника углерода. Таким образом, на определенных этапах процесса синтеза различные исходные источники углерода превращаются в одинаковые соединения. Очень возможно, что эти исходные соединения углерода превращаются в одинаковые промежуточные продукты обмена веществ еще до начала синтеза (Капутто, 1949).
Промежуточные продукты обмена веществ должны, без сомнения, служить возможным источником углерода для гриба, который способен их образовывать. Если гриб превращает соединение А в соединение Б, то это последнее также должно служить возможным источником углерода для данного гриба. Неспособность гриба усваивать соединение Б указывает на то, что это соединение не является в действительности промежуточным продуктом обмена веществ. Однако в этой простой гипотезе не учитываются две следующие возможности: во-первых, соединение Б может проникать в клетку гриба не столь легко, как соединение А, и, во-вторых, соединение Б в испытываемых концентрациях может быть ядовито для гриба.
Нередко промежуточные продукты диссимиляции сахаров поступают в среду, из которой их можно затем выделить. Так, наличие ацетальдегида устанавливают, добавляя к среде бисульфит. Ацетальдегид образует с бисульфитом нерастворимое соединение. Иногда накопление определенного промежуточного продукта в среде обуславливается замедленным протеканием следующего этапа обмена веществ. В этих случаях промежуточные продукты обмена веществ, подвергаясь дальнейшим превращениям, со временем исчезают из среды. К таким промежуточным продуктам обмена веществ относятся ацетальдегид, этиловый спирт и пировиноградная кислота.
Нельзя утверждать, что усвоение сахара или какого-либо иного источника углерода происходит одинаково у всех видов грибов. Нет также никаких оснований предполагать, что у данного вида гриба существует лишь один путь усвоения углеводов. Nord F.F. и Mull R.P. (1945) считают, что диссимиляция углеводов различными видами Fusarium может происходить в результате окисления, в результате разрыва углеродной цепочки или, наконец, с участием реакций фосфорилирования. Относительная значимость этих трех процессов диссимиляции сахаров зависит от вида гриба и от внешних условий. Образование двумя различными видами грибов одинакового промежуточного продукта обмена веществ не является еще доказательством того, что в обоих случаях механизм образования этого продукта одинаков. Так, дрожжи и некоторые виды Fusarium образуют спирт, однако биохимические процессы, в результате которых эти микроорганизмы превращают глюкозу в спирт, различны (Лилли, 1957).
Похожие работы
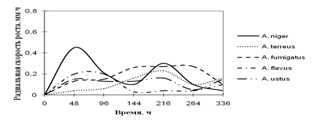
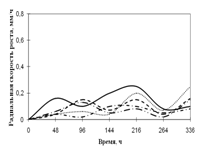
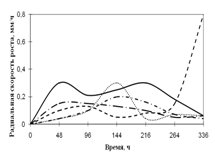
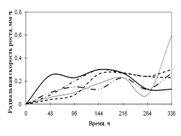
... просмотренных. Наблюдение проводят на 1, 2, 3, 7, 15-е сутки. Стекла микроскопировали, учитывая прорастание спор (%), длину проростков (в мкм) (Кураков, 2001). ГЛАВА 3. ИССЛЕДОВАНИЕ РОСТА МИКРОМИЦЕТОВ НА РАЗЛИЧНЫХ СУБСТРАТАХ Объектами исследования явились 10 штаммов коллекционных микроскоскопических грибов родов Aspergillus: A. niger, A. ustus, A. terreus, A. flavus, A. fumigatus, а также ...
... 5 видов разлагали хитин, 8 – выделяли тирозиназу и лакказу (Parton, 1988). Отмечают высокую целюлозолитическую активность у штаммов видов Trichoderma, Aspergillus niger и Penicillium miszynsky, выделенных из водных местообитаний (Hopper, 1995). Микроскопические грибы могут прямо или косвенно влиять на активность и развитие других гидробионтов. Пути воздействия могут быть различными – от прямого ...
... города Чернигова мною было собрано 13 видов дереворазрушающих грибов. Эти грибы были собраны в 3 – х географических пунктах окрестностей города, а именно Массаны, Яцево, район р. Десна. Распространение трутовиковых грибов в природных условиях определяется рядом факторов, важнейшим из которых служит субстрат, т.е. соответствующая древесина. Способность некоторых видов развиваться только на живых ...
... к ассимиляции ЭДТА и глюкозы в процессе длительного культивирования с добавлением субстрата В предыдущем разделе было показано, что ассимиляция глюкозы бактериальным штаммом LPM-4 индуцируется в процессе деградации ЭДТА, а кометаболизм ЭДТА и глюкозы у штамма LPM-4 не оказывает влияния на деградацию ЭДТА. В данном разделе целью работы было исследование: 1) сохраняется ли способность клеток ...
0 комментариев