Навигация
Экзоцитоз нейромедиаторов
68465
знаков
0
таблиц
5
изображений
2. Экзоцитоз нейромедиаторов
В нейрохимическом плане лучше других синапсов изучен электромоторный синапс электрического органа рыб, где нейро-медиатором служит АХ. В начале 70‑х годов в лаборатории В. Уит-такера в ФРГ впервые удалось выделить изолированную фракцию синаптических пузырьков из электрического органа ската Torpedo marmorata. Именно на этом объекте с помощью биохимических, иммуноцитохимических методов и ядерного магнитного резонанса получены фундаментальные сведения о структуре и функциях синаптических пузырьков и разработана схема их жизненного цикла.
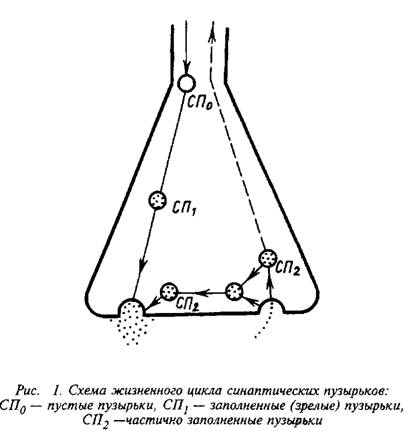
В аппарате Гольджи сомы нейрона формируются мембранные образования в виде пузырьков, не заполненных медиатором. Эти пузырьки направляются в пресинаптическое окончание с помощью системы быстрого аксонного транспорта. В пресинаптическом окончании пузырьки заполняются медиаторами посредством АТФ-зависимого протонного насоса. Молекулы протонной АТФазы входят в состав мембраны синаптических пузырьков и поддерживают определенный уровень мембранного потенциала. Мембрана синаптического пузырька содержит также стимулируемую кальмодулином Са+-АТФазу, которая обеспечивает поглощение пузырьками ионов Са. Популяция зрелых пузырьков. Циклы экзоцитоз – эндоцитоз повторяются.
Эта схема согласуется с электрофизиологическими данными о квантовом характере секреции нейромедиатора и о численности квантов разных размеров в одном и том же пресинаптическом окончании, а также с радиохимическими сведениями о предпочтительном освобождении вновь синтезированного медиатора. Таким образом, пресинаптическос окончание можно рассматривать как систему, в определенной мере автономную по отношению к телу нейрона.
Синаптические пузырьки диаметром 50–60 нм, так называемые малые прозрачные синаптические пузырьки, аналогичные холинергическим синаптическим пузырькам из электрического организма ската, выделены из разных отделов нервной системы представителей практически всех таксономических групп животных. Эти пузырьки отличаются низкой электронной плотностью содержимого. Они заполнены низкомолекулярными нейромедиаторами в отличие от больших электронно-плотных пузырьков, заполненных медиаторами пептидной природы.
Ключевую проблему в изучении экзоцитоза нейромедиаторов представляет вопрос о механизмах сближения синаптического пузырька с активной зоной пресинаптической мембраны и взаимодействия мембраны пузырька с активной зоной. Имеются данные о том, что эти процессы зависят. от Са+ – универсального внутриклеточного посредника, участие которого в секреторных процессах может быть обусловлено активацией актомиозино-вых филаментов цитоскелета, мембранной фосфолипазы А2, аде-нилатциклазы, Са+/кальмодулин-зависимых протеинкиназ и ряда других Са+-связывающих белков.
Известно, что связывание Са+ с кальмодулином индуцирует фосфорилирование ряда белков синаптосом. Ингибиторы кальмодулина и кальмодулинкиназы блокируют освобождение нейромедиаторов, вызываемое деполяризацией синаптосом. При воздействии различных факторов, влияющих на количество освобождаемого медиатора и на фосфорилирование белков, выявлена корреляция между изменениями секреции нейромедиаторов и фосфорилированием белков синаптосом.
Независимо от того, из какого отдела нервной системы они получены и какой нейромедиатор содержат, малые синаптические пузырьки характеризуются специфическим набором интегральных мембранных белков, к которым относятся синапсины – фосфопротеины, фосфорилируемые цАМФ- и Са+-зависимыми протеинкиназами, синаптофизин – гликопротеин, пронизывающий мембрану; синаптобревин – негликозилированный белок, находящийся на цитоплазматической поверхности пузырьков; белок SNAP‑25; синтаксин, синаптогамин, синаптопорин и др.
Особая роль в сближении синаптического пузырька с активной зоной отводится синапсину. Этот белок, который состоит из двух полипептидов с молекулярной массой 86 и 80 кД, ассоциирован с цитоплазматической поверхностью мембраны синаптического пузырька. При микроинъекции фосфорилированной формы синапсинов в пресинаптическое окончание гигантского аксона кальмара наблюдается повышение амплитуды и скорости нарастания постсинаптического потенциала, что свидетельствует об увеличении секреции медиатора; дефосфорилированные формы синапсиноз не вызывали такого эффекта.
Аналогичное увеличение секреции медиатора происходит при микроинъекции Са+/кальмодулина. Показана способность очищенных синапсинов взаимодействовать в зависимости от состояния фосфорилирования с белками мембраны синаптического пузырька и с F‑актином цитоскелета пресинаптического окончания. Предложена следующая схема участия синапсинов в экзоцитозе. В отсутствии деполяризации пресинаптического окончания, когда концентрация Са+ в цитоплазме низка, дефосфорилированный синапсин, связанный с цитоплазматической поверхностью пузырька, взаимодействует с цитоскелетом, обеспечивая резервирование и иммобилизацию пузырька. При деполяризации пресинаптической мембраны происходит вход Са+ в пресинаптическое окончание - активация Са+/кальмодулинкиназы -> фосфорилирование синапсина I -» ослабление связи между синапсином и пузырьком, а также синапсином и F‑актином. В результате синаптический пузырек перемещается вдоль микротрубочек на стратегическую позицию v активной зоны.
Далее наступает цепь реакций, обеспечивающих контакт пузырька с пресинаптической мембраной и его плавление. Здесь опять-таки процесс инициируется Са+, который связывается с другим белком пузырька синаптогамином Именно Са+-синаптогамин взаимодействует с фосфолипидами и с комплексом других белков, регулирующих плавление везикулы, – синаптобревином, синтаксыном и синаптофизином. В заключение происходит активация белка синаптопорина, формирующего пору, какал, через который изливается содержимое везикулы.
В поисках молекулярных механизмов слияния мембраны синаптического пузырька с пресинаптической мембраной выявлено, что ботулинический и столбнячный токсины, блокирующий экзоцитоз нейромедиаторов из синаптических пузырьков, повреждают именно указанную выше триаду белков пузырька – синаптобревин, синтаксин и SNAP‑25.
Ряд других деталей конечного этапа экзоцитоза пока не выяснен. Существует предположение, что выброс нейромедиаторов происходит при активном сокращении стенок пузырька с участием актомиозинподобных белков, активируемых ионами Са.
Синаптическая пластичность означает способность синапсов к функциональным и морфологическим перестройкам в процессе синаптической активности. Свойство пластичности синапсов составляет основу таких явлений, как обучение, память.
Изменение эффективности синапса после активации определяется увеличением или уменьшением амплитуды постсинаптических потенциалов, которые в свою очередь связаны с изменением количества нейромедиатора, высвобождаемого из пресинаптических окончаний.
Существует несколько основных фаз постактивационных изменений синаптической эффективности.
1. Облегчение – повышение амплитуды постсинаптического потенциала в начальный период ритмической серии пресинаптических импульсов.
2. Кратковременная посттетаническая потенциация – повышение амплитуды постсинаптического потенциала при ритмической активации нервного окончания в течение десятков секунд.
3. Постактивационная депрессия постсинаптических потенциалов, которая развивается параллельно и взаимодействует с процессом потенциации.
4. Длительная потенциация постсинаптических потенциалов, которая медленно развивается вслед за кратковременной потенциацией и продолжается в течение часов и даже дней.
Эти явления связаны с изменением концентрации Са+ в пресинаптических окончаниях во время ритмической активности. Длительные модификации синаптической эффективности ассоциируются с изменениями фосфорилирования синаптических белков.
Следует упомянуть, что наряду с пресинаптическими механизмами существуют и постсинаптические механизмы, основанные на изменениях чувствительности рецепторов нейромедиаторов.
Похожие работы

... В этом случае раздражения разделены интервалом времени, а не пространством и более высокую эффективность повторного раздражения можно считать результатом временной суммации. В современных учебниках по физиологии приводятся понятия временной и пространственной суммации, но теперь об их механизме известно больше, чем в то время, когда они были впервые описаны. Нервные волокна многократно ветвятся, ...
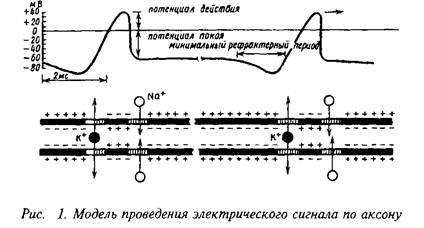
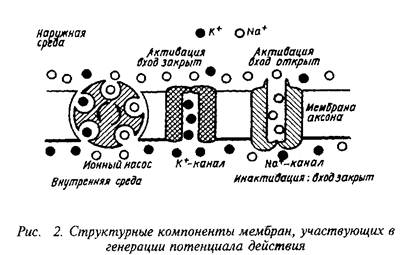
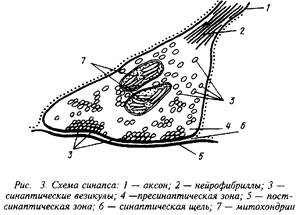
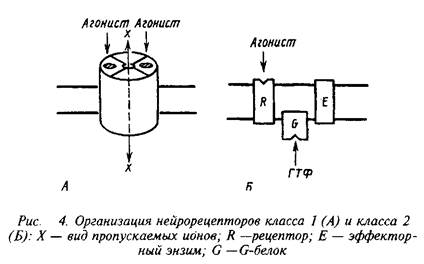
... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
... специфических сигналов. Представление о синапсах связано с Шеррингтоном (1935), высказавшим предположение о существовании специальных структурно-функциональных образований, обеспечивающих контакты между нейронами. Особенности рефлекторных реакций и некоторые свойства нервных центров обусловлены процессами, происходящими на синапсах. Синапс включает в себя три компонента: пресинаптический, ...
... в глутамат с образованием янтарного полуальдегида. HOOC-CH2-CH2-CH2-NH2 à HOOC-CH2-CH2-CH(O) ГАМК Янтарный полуальдегид ГАМК в организме γ–аминомасляная кислота является основным ингибиторным трансмиттером (передатчиком) в ЦНС. Она осуществляет нейрональную трансмиссию в 1/3 всех синапсов головного и спинного мозга. ГАМК играет важную роль в регуляции двигательных функций. При этом ...
0 комментариев