Навигация
3.5 Сопутствующие медиаторы
Сопутствующие, или сосуществующие, медиаторы – это синоптические посредники, которые характеризуются прежде всего совместной локализацией и совместным высвобождением. Под совместной локализацией имеется в виду синтез и депонирование медиаторов в одном и том же нейроне, их присутствие в одних и тех же пресинаптических окончаниях, но не обязательно в одних и тех же синаптических пузырьках. Так, низкомолекулярные классические нейромедиаторы депонируются преимущественно в мелких оптически прозрачных пузырьках, а пептидные медиаторы – в крупных оптически плотных пузырьках, хотя имеются данные и о случаях локализации этих двух видов медиаторов в одних и тех же оптически плотных пузырьках. Различие в системах депонирования этих двух видах медиаторов обусловлено различиями мест их синтеза: классические нейромедиаторы синтезируются в цитоплазме пресинаптических окончаний и затем поступают в синаптические пузырьки, а пептидные медиаторы синтезируются в аппарате Гольджи, т.е. в соме нейрона, и доставляются в нервные окончания уже упакованными в пузырьки.
Под совместным высвобождением понимается экзоиитоз двух медиаторов в результате одного и того же процесса активации пресинаптического окончания, хотя под процессом активации в данном случае подразумевается не одиночный пресинаптический потенциал действия, а разряд потенциалов действия с той или иной частотой. Еще один признак сопутствующих медиаторов состоит в том, что они вызывают функциональные изменения в одной и той же клетке-мишени.
Иммуноцитохимическими методами в центральных и периферических нейронах прослежены разнообразные виды сочетаний представителей медиаторных групп: 1) несколько классических нейромедиаторов; 2) классический) нейромедиатор + нейропетид несколько нейропептидов, имеющих общую молекулу-предшественник; 4) несколько нейропептидов, кодируемых разными генами. К этим сочетаниям могут добавляться пурины.
3.6 Локализация нейромедиаторных путей
Прежде чем рассматривать наиболее изученные индивидуальные синаптические системы и соответствующие медиаторы, целесообразно рассмотреть общую картину локализации нейромедиаторных путей в мозге, отметив одновременно самые общие характеристики функций соответствующих систем.
Химические синапсы распределены в нервной ткани не в случайном порядке, а организованы в определенных группах нейронов. Для того чтобы уверенно картировать какие-либо меди аторные пути в головной мозге, необходимо иметь доступные и надежные специфические маркеры, с помощью которых можно визуализировать интересующие исследователя межклеточные взаимодействия.
Существуют три основных методических подхода для решения этой задачи. Первый способ – избирательное окрашивание нейронов, выделяющих определенный нейромедиатор, может осуществляться с помощью преобразования естественного медиатора в его флуоресцирующее производное. В этом случае флуоресценция определенных групп клеток поможет выявить специфические связи в структурах мозга. Второй экспериментальный подход связан с введением молекул медиатора, предварительно меченного радиоактивным изотопом. Нейронные окончания, содержащие исследуемый медиатор, способны избирательно захватывать метку. Затем их легко выявить методом авторадиографии. Третий способ обнаружения специфических связей в нервной системе состоит в использовании высоко специфичной способности узнавать либо антигенные детерминанты медиатора, либо определенные ферментные белки, участвующие в метаболизме нейромедиаторов, либо нейрорецептор-ные компоненты на мембране клетки. Последние считаются наиболее убедительным свидетельством в пользу существования конкретных нейрохимических взаимодействий между клетками и зонами мозга. Обычно для иммунохимической идентификации используют флуоресцентный краситель или изотоп, который маркирует антитела. В последние годы широко распространились методы, использующие антитела, меченные частицами тяжелых металлов, например коллоидного золота, железа и др.
Указанные способы позволили получить весьма ценную информацию о детальном анатомическом распределении различных синапсов, В первую очередь это помогло локализовать катехоламинергические синапсы, а также синапсы, содержащие ферментные системы с уникальными метаболитами, такими как ГАМК. В отличие от катехоламинов аминокислотные нейромедиаторы – L‑глутамат, L‑аспартат, глицин – участвуют в метаболических процессах практически всех клеток, в связи с чем выбор специфических маркеров для соответсвующих синапсов затруднен. В этих случаях обнаружение самого нейромедиатора или его ферментных систем еще не позволяет судить о природе химического синапса и тогда использует косвенный подход, например измерение высвобождаемых медиаторных аминокислот из нервных окончаний при их разнообразной стимуляции.
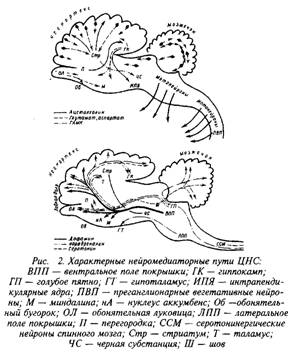
Общая схема рапределения нейромедиаторных связей по структурам представлена на рис. 2. Как видно из схемы, пока имеется недостаточно фактов, чтобы представить полную картину химического картирования мозга. Это требует дальнейшей большой совместной работы морфологов и нейрохимиков. Однако для ряда путей, особенно функционирующих на основе биогенных аминов, эти данные уже получены. Они локализованы преимущественно в нейронах, входящих в состав полосатого тела, вентролатерального ядра, черного вещества и голубого пятна. Аксоны этих нейронов проецируются, как правило, в гипоталамус, мозжечок, передний мозг.
Полагают, что моноаминовые нейромедиаторные пути имеют отношение к проявлению эмоций, регуляции настроения, поддержанию состояния бодрствования и др. Нарушение обмена норадреналина связывают с возникновением ряда психоэмоциональных расстройств. Воздействие на указанные пути фармакологическими средствами, компенсирующими дефицит или избыток нейромедиатора, способствует в ряде случаев снижению симптоматики шизофрении, маниакально-депрессивных состояний и др.
Нейроны, содержащие дофамин, сосредоточены в областях среднего мозга, особенно их много в черной субстанции и вентролатеральной покрышке. Многие из этих нейронов посылают свои аксоны в передний мозг, где они участвуют в развитии эмоциональных реакций. Важную роль дофаминовая медиаторная система играет в регуляции сложных двигательных функций.
Обнаружено, что нарушения дофаминергических путей приводит к затруднению движений, особенно стереотипных, к возникновению непроизвольного дрожания и скованности мышц, т.е. к появлению характерных симптомов паркинсонизма. Фармакологические препараты, содержащие метаболиты дофамина, проникающие в головной мозг, способны смягчить проявление болезни. Гиперфункция дофаминергической системы связана с механизмами шизофрении.
Моноаминовый нейромедиатор серотонин сосредоточен в области ствола мозга, где находятся так называемые «ядра шва». Нейроны этого центра проецируются в гипоталамус, таламус и другие области мозга.
Как полагают, они участвуют в терморегуляции, сенсорном восприятии и процессах сна.
Лекарственные средства, которые способны частично удалить из синапсов моноаминовые нейромедиаторы, вызывают депрессию, тогда как все препараты, применяемые для лечения клинической депрессии, обычно повышают содержание этих нейромедиаторов, усиливают их действие.
Ацетилхолин широко представлен в разных отделах нервной системы, основное его количество находится в периферических нервно-мышечных синапсах, рецепторы которых относятся к категории так называемых никотиновых. В ЦНС ацетилхолин сосредоточен. преимущественно в базальных ганглиях, таламусе и сером веществе. Соответствующие рецепторы в мозге относятся главным образом к категории мускариновых. Вставочные холинергические нейроны обнаружены в хвостатом ядре, переднем роге латерального желудочка, которые являются одними из наиболее богатых ацетилхолиновых мозговых структур.
Полагают, что АХ в подкорковых структурах участвует в тонкой регуляции сложных двигательных функций, в частности в механизмах инициации движения, двигательных стереотипах и др. Поражение холинергической иннервации в структурах мозга сопровождается нарушением и извращением двигательных функций. Так, например, при паркинсонизме наряду с нарушением дофаминергической трансмиссии отмечается гиперактивность некоторых холинергических систем мозга. Поэтому для лечения этого заболевания используют препараты, содержащие холинолитики, которые снижают уровень ацетилхолина или стимулируют работу ацетилхолинэстеразы. Эти вещества вводят совместно с аналогами L‑ДОФА для компенсации дефицита дофаминергических путей. Нарушение холинергической иннервации характерно и для ряда сенильных болезней мозга.
Поражение холинергической передачи в периферической нервной системе, в частности нервно-мышечных синапсах, связано с симптомами «усталости» или «слабости» мышц. Полагают, что в основе тяжелого заболевания – миастении гравис – лежит аутоиммунный процесс. Организм вырабатывает аутоантитела, которые блокируют функцию холинорецепторов.
Наиболее широко распространеными в ткани мозга рецепторами и, соответственно, нейромедиаторами являются некоторые аминокислоты. Центральное место среди них занимает L‑глутаминовая кислота – основной возбуждающий нейромедиатор. Глутаматергические синапсы распространены в коре головного мозга, гиппокампе, полосатом теле и гипоталамусе. Нисходящие глутаматергические пути обнаружены практически во всех структурах головного мозга, проекции которых идут от коры к подкорковым структурам. Выявление глутаматергических связей в головном мозге проводится преимущественно методом физиологической идентификации по. высвобождению нейромедиатора. В последние годы на основе изучения структуры и свойств глутаматных рецепторов появилась возможность визуализации нейрорецепторов глутамата с помощью моноклональных и политональных антител,
Нейрорецепторы глутамата располагаются кластерами на постсинаптической мембране. Выявление глутаматных рецепторов на мембране клеток с помощью иммуногистохимических методов является более надежным способом идентификации глутаматергических связей по сравнению с другими методами.
Нарушение глутаматергической медиации связано с целым рядом патологических состояний нервной системы: эпилепсией, расстройствами вестибулярной системы, ишемическими проявлениями и др. Глутаминовая кислота и некоторые ее аналоги используются в качестве терапевтического лекарственного средства при хронической недостаточности аминокислотного обмена, вегетососудистой дистонии и эпилепсии.
Присутствие в разных структурах мозга ГАМК – первого по значимости тормозного нейромедиатора – показано методами авторадиографии. Топографическое распределение самого радиоактивно меченного нейромедиатора или образующего его фермента – глутаматдекарбоксилазы – в головном мозге неравномерно. К областям, содержащим наиболее высокую концентрацию ГАМК, относятся черное вещество, бледный шар, гипоталамус и мозжечок. Аминокислота содержится преимущественно в сером веществе головного и спинного мозга.
Данные о функциональной роли ГАМК-ергической передачи в головном и спинном мозге постоянно обогащаются новыми фактами. Она принимает участие в регуляции моторной активности, поддержании судорожного порога, формировании эмоционального поведения. ГАМК-ергическая система участвует в осуществлении условных рефлексов, организации процессов обучения и памяти у млекопитающих. При этом она тесно взаимодействует с другими медиаторными системами мозга: дофаминергической, холинергической и глутаматергической.
Имеется очень большое количество данных о вовлечении системы ГАМК в механизмы многих метаболических расстройств нервной системы. Установлено, что нарушения этой системы связаны с прявлениями эпилепсии, хореи Гентингтона, паркинсонизма и некотрых других поражений экстрапирамидной системы. При терапевтическом применении соединений, содержащих эту аминокислоту или ее аналоги, обнаруживаются позитивные клинические эффекты. Имеются данные о благоприятном влиянии производных ГАМК на больных эпилепсией, хореей Гентингтона и паркинсонизмом. Эти же препараты способны усиливать дыхание, энергетический обмен нервной ткани, улучшать показатели мозгового кровообращения и метаболизма глюкозы.
Как упоминалось, методология гистохимического исследования локализации ГАМК или образующего ее фермента – глу-таматдекарбоксилазы – неприменима для выявления других тормозных систем – глицина и таурина и их рецепторов. Анализ их распределения производят, как правило, с использованием методов электрофизиологической регистрации высвобождения нейропередатчика из нервных окончаний при их разнообразной стимуляции.
Глицин и его рецепторы локализованы в зонах моста, продолговатого мозга и серого вещества спинного мозга, включая передние и задние рога. Авторадиографически была установлена локализация участков высокоаффинного захвата глицина преимущественно в аксо-аксональных и аксодевдритных синапсах. Скопление гранул отмечены также вокруг клеточных тел спинальных мотонейронов. У больных с некоторыми врожденными метаболическими аномалиями, связанными с повышением содержания глицина в ткани мозга и крови, может развиваться гиперглицинемия, которая сопровождается симптомами нарушения некоторых психоэмоциональных функций. Полагают, что такие расстройства могут быть следствием поражения обычных путей деградации глицина в нервной клетке.
Интересные данные были получены для таурина. Уровень таурина в разных зонах мозга оказался практически одинаковым, за исключением следующих структур: медиального коленчатого тела, гипофиза и шишковидной железы. Общая концентрация таурина и цистеинсульфонатдекарбоксилазы в спинном мозге и таламических ядрах совпадает с количеством ГАМК в этих структурах, однако локализация их по зонам внутри структур существенно различается. Все исследователи таурина сходятся во мнении о его необычайно высоком содержании в коре мозжечка, которое почти в 5 раз превышает уровень ГАМК в этой структуре. Показано, что таурин локализуется преимущественно в звездчатых нейронах молекулярного слоя. Это позволило предположить существование тауринергических нейронов. Вместе с тем авторадиографическое изучение распределения таурина свидетельствует и о его преимущественно глиальной локализации.
Существует достаточно веские аргументы в пользу того, что таурин является также важным компонентом питания живых организмов, так как он не синтезируется у млекопитающих, включая человека. Клинически тауриновый дефицит может выражаться в эпилептических припадках, наследственной атаксии Фридрейха, зрительной дисфункции, называемой в просторечии «куриной слепотой», и др. Широко обсуждается возможность участия таурина в патогенезе судорожно-пароксизмальных состояний. Выяснилось, что таурин, введеный в желудочки мозга крысы, подверженной судорогам, является более мощным, чем ГАМК, противосудорожным агентом. Однако в клинической практике таурин не проявляет стабильных противосудорожных эффектов и пока не нашел широко применения.
Похожие работы

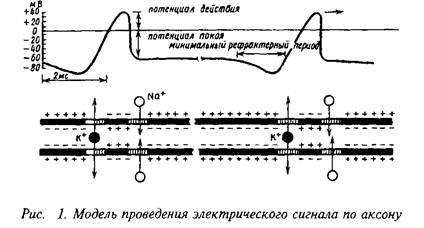
... В этом случае раздражения разделены интервалом времени, а не пространством и более высокую эффективность повторного раздражения можно считать результатом временной суммации. В современных учебниках по физиологии приводятся понятия временной и пространственной суммации, но теперь об их механизме известно больше, чем в то время, когда они были впервые описаны. Нервные волокна многократно ветвятся, ...
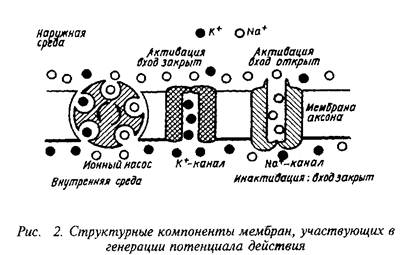
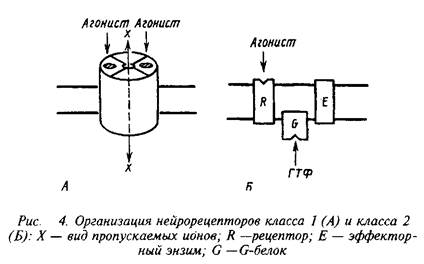
... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
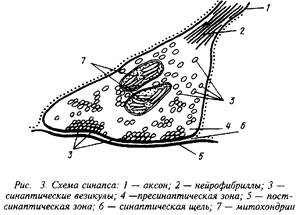
... специфических сигналов. Представление о синапсах связано с Шеррингтоном (1935), высказавшим предположение о существовании специальных структурно-функциональных образований, обеспечивающих контакты между нейронами. Особенности рефлекторных реакций и некоторые свойства нервных центров обусловлены процессами, происходящими на синапсах. Синапс включает в себя три компонента: пресинаптический, ...
... в глутамат с образованием янтарного полуальдегида. HOOC-CH2-CH2-CH2-NH2 à HOOC-CH2-CH2-CH(O) ГАМК Янтарный полуальдегид ГАМК в организме γ–аминомасляная кислота является основным ингибиторным трансмиттером (передатчиком) в ЦНС. Она осуществляет нейрональную трансмиссию в 1/3 всех синапсов головного и спинного мозга. ГАМК играет важную роль в регуляции двигательных функций. При этом ...
0 комментариев