Навигация
Ограничения закона Харди-Вайнберга
41569
знаков
1
таблица
0
изображений
3. Ограничения закона Харди-Вайнберга
3.1 Идеальные условия для закона
В полной мере закон Харди–Вайнберга применим к «идеальной популяции», которая характеризуется следующими признаками:
· бесконечно большие размеры;
· неограниченная панмиксия;
· отсутствие мутаций;
· отсутствие иммиграции особей из соседних популяций;
· отсутствие естественного отбора.
В природных популяциях ни одно из этих условий не соблюдается, поэтому и закон Харди-Вайнберга носит условный характер. Тем не менее он реально отражает тенденции в характере распределения частот тех или иных аллелей и генотипов.
3.2 Частоты аллелей (Верн Грант)Генофонд локальной популяции обычно содержит помимо мономорфных различные полиморфные гены. В каждом данном поколении аллельные формы полиморфных генов представлены с некоторой определённой частотой. Так, например, ген А, имеющий два аллеля, А и а, может быть представлен в генофонде одного поколения в соотношении 70% аллелей А и 30% аллелей а. Каковы в таком случае будут ожидаемые частоты аллелей в следующем поколении? [3]
В популяции диплоидного организма эти аллели содержатся в гомозиготных и гетерозиготных генотипах АА, аа и Аа, которые будут встречаться в определённых соотношениях в любом данном поколении. Они служат родительскими генотипами для следующего поколения. В связи с этим возникает вопрос: каковы ожидаемые соотношения генотипов во втором и в последующих поколениях?
Ожидаемые частоты аллелей и генотипов можно определить по закону Харди — Вайнберга. Этот закон действует при следующих условиях. Предполагается, что популяция достаточно велика, для того чтобы ошибки выборки не оказывали существенного влияния на частоты в последовательных поколениях. Популяция изолирована, иммиграция отсутствует, составляющие популяцию особи вносят равное число функционирующих гамет; иными словами, разные генотипы размножаются одинаково успешно. И наконец, предполагается, что в популяции преобладает случайное скрещивание. Случайное скрещивание, или панмиксию, можно с равным успехом определять в терминах особей или в терминах гамет. Если иметь в виду особей, то случайное скрещивание происходит в тех случаях, когда особи с различной генетической конституцией скрещиваются независимо от своих генотипов. Например, самка с генотипом АА может скрещиваться с самцами АА, Аа или аа, не проявляя никакого предпочтения к самцам какого-то одного типа.
Панмиксию можно определить точнее, если исходить из наличия в гаметном фонде множества гамет. В этом смысле случайное скрещивание означает, что любая женская гамета с одинаковой вероятностью может быть оплодотворена мужской гаметой любого типа и что эта вероятность прямо пропорциональна частоте мужских гамет данного типа в гаметном фонде. Короче говоря, гаметы, несущие разные аллели, соединяются в пары пропорционально их относительным частотам в гаметном фонде. Особи, составляющие популяцию в каждом данном поколении, представляют собой в таком случае произведения разных пар гамет, случайно извлеченных из гаметного фонда предшествующего поколения.
В популяции, соответствующей указанным выше условиям, согласно закону Харди — Вайнберга, частоты аллелей будут оставаться постоянными из поколения в поколение, и при случайном скрещивании в одном поколении генотипы достигнут равновесных частот, которые сохранятся в дальнейшем. Например, закон постоянства частот аллелей мы проиллюстрируем количественным примером. Допустим, что популяция некоего диплоидного вида, полиморфного по гену А, в исходном поколении содержит разные генотипы в следующем соотношении: 60% АА, 20% Аа и 20% аа. Проследим за аллелями А на протяжении двух поколений.
1. Частоты аллелей в первом поколении. Поскольку частоты генотипов заданы как 0.60 AA + 0.20Aa + 0.20 aa, частоты аллелей (q) в этом поколении должны составлять qA = (0.60 + 0.60 + 0.20) \ 2 = 0.70, а qа = (0.20 + 0.20 + 0.20) \ 2 = 0.30.
2. Гаметный фонд первого поколения. Предполагается, что все особи одинаково плодовиты, поэтому диплоидные особи будут производить гаплоидные гаметы в соотношении 70% А и 30% а. Частоты аллелей в гаметном фонде такие же, как и в исходном генофонде. [3]
3. Случайное скрещивание. Гаметы для образования зигот второго поколения извлекаются из фонда случайным образом; при этом возможны также попарные сочетания;
| Женские гаметы | Мужские гаметы | |
| 0.70 A | × | 0.70 A |
| 0.70 A | × | 0.30 a |
| 0.30 a | × | 0.70 A |
| 0.30 a | × | 0.30 a |
4. Частоты зигот во втором поколении. Приведенная выше система свободного скрещивания даёт следующие результаты:
0.49 АА; 0.21 + 0.21 = 0.42 Аа; 0.09 аа.
Считается, что все зиготы обладают одинаковой жизнеспособностью; следовательно, приведённые цифры дают ожидаемые равновесные частоты генотипов во втором поколении.
Можно заметить, что данная популяция не находилась в равновесии в отношении частот генотипов в первом поколении, но достигла равновесного состояния в результате свободного скрещивания всего лишь в одном поколении;
5. Частоты аллелей во втором поколении. Генофонд второго поколения, очевидно, будет содержать два аллеля со следующими частотами:
А = ( 0.49 + 0.49 + 0.42) \ 2 = 0.70, а = (0.42 + 0.09 + 0.09) \ 2 = 0.30.
Таким образом, частоты аллелей во втором поколении такие же, какими они были в первом поколении. [3]
Эффекты инбридинга. Инбридинг – это близкородственное скрещивание, скрещивание организмов, имеющих общего предка. При инбридинге повышается вероятность наличия у скрещиваемых организмов одних и тех же аллелей. В большой популяции при условии случайного скрещивания частоты генотипов быстро достигают равновесного состояния, которое сохраняется в дальнейшем. Нарушения случайности скрещивания вызывают отклонения частот генотипов от равновесия Харди — Вайнберга.Допустим, что некая популяция в нулевом поколении состоит исключительно из гетерозигот Аа. Размножение происходит путем самооплодотворения. В первом поколении популяция будет иметь следующий состав: 25% АА, 50% Аа и 25% аа. В последующих поколениях сокращение класса гетерозигот будет продолжаться с равномерной скоростью. После самооплодотворения в семи поколениях (т. е. в 7-м инбредном поколении) популяция будет содержать почти 50% АА и почти 50% аа. [3]
3.3 Мутационный процесс Организмы, обладающие удачными вариантами признаков, имеют большую вероятность по сравнению с другими организмами выжить и оставить потомство. Вследствие этого полезные вариации в ряду поколений будут накапливаться, а вредные или менее полезные вытесняться, элиминироваться. Это и называется процессом естественного отбора, который играет ведущую роль в определении направления и скорости эволюции.
Прямая взаимосвязь между степенью генетической изменчивости в популяции и скоростью эволюции под действием естественного отбора была доказана математическим путем Р. Фишером [1930] в его фундаментальной теореме естественного отбора. Фишер ввел понятие приспособленности и доказал, что скорость возрастания приспособленности популяции в любой момент времени равна генетической вариансе приспособленности в тот же момент времени. Однако прямые доказательства этого факта были получены лишь в конце 60-х годов ХХ столетия.
Мутационный процесс служит источником появления новых мутантных аллелей и перестроек генетического материала. Однако возрастание их частоты в популяции под действием мутационного давления происходит крайне медленно, даже в эволюционном масштабе. К тому же подавляющее большинство возникающих мутаций устраняются из популяции в течение немногих поколений уже в силу случайных причин. Неизбежность такого течения событий впервые обосновал Р. Фишер в 1930 году. Для человека и других многоклеточных показано, что мутации обычно возникают с частотой от 1 на 100 000 до 1 на 1 000 000 гамет. Новые мутанты, хотя и довольно редко, но постоянно появляются в природе, поскольку существует множество особей каждого вида и множество локусов в генотипе любого организма. Например, число особей того или иного вида насекомых обычно составляет около 100 млн. (108). Если предположить, что средняя мутабельность по одному локусу равна 1 мутации на 100 000 (10-5) гамет, то среднее число вновь возникающих в каждом поколении мутантов по этому локусу для данного вида насекомых составит ![]() . (Частота возникновения мутаций умножается на число особей и еще на два, так как любая особь представляет собой продукт слияния двух гамет.) В генотипе человека имеется около 100 000 (105) локусов. Предположим, что у человека темп мутирования такой же, как у дрозофилы; в этом случае вероятность того, что генотип каждого человека содержит новый аллель, отсутствовавший в генотипе его родителей, равна
. (Частота возникновения мутаций умножается на число особей и еще на два, так как любая особь представляет собой продукт слияния двух гамет.) В генотипе человека имеется около 100 000 (105) локусов. Предположим, что у человека темп мутирования такой же, как у дрозофилы; в этом случае вероятность того, что генотип каждого человека содержит новый аллель, отсутствовавший в генотипе его родителей, равна ![]() . Иными словами, каждый человек в среднем несет около двух новых мутаций.
. Иными словами, каждый человек в среднем несет около двух новых мутаций.
Проделанные выше расчеты основаны на частотах возникновения мутаций, обладающих внешним проявлением. В целом по геному темп мутирования составляет не менее ![]() замен на одну нуклеотидную пару в год. У млекопитающих число нуклеотидных пар в диплоидном геноме составляет около
замен на одну нуклеотидную пару в год. У млекопитающих число нуклеотидных пар в диплоидном геноме составляет около ![]() . Следовательно, нуклеотидные замены у млекопитающих происходят с частотой не менее
. Следовательно, нуклеотидные замены у млекопитающих происходят с частотой не менее ![]() в год на диплоидный геном. Ясно, что мутационный процесс обладает колоссальными возможностями поставлять новый наследственный материал.
в год на диплоидный геном. Ясно, что мутационный процесс обладает колоссальными возможностями поставлять новый наследственный материал.
Важный шаг в генетике популяций был сделан в 1926 году С. С. Четвериковым. Исходя из закона Харди – Вайнберга, С. С. Четвериков доказал неизбежность генетической разнородности природных популяций при том, что новые мутации непрерывно появляются, но остаются обычно скрытыми (рецессивными), а в популяции идет свободное скрещивание.
Из расчетов Четверикова следовало, а впоследствии это было полностью подтверждено практикой, что даже редкие и вредные для особи мутантные гены будут надежно укрыты от очищающего действия естественного отбора в гетерозиготах (организмах со смешанной наследственностью) с доминирующими безвредными генами нормального дикого типа. Это значит, что даже вредная гетерозигота (организм с однородной наследственностью) мутация будет сохраняться в виде генетической «примеси» в течение ряда поколений. Мутация будет как бы поглощена популяцией, из-за чего за внешним однообразием особей одной популяции неизбежно скрывается их огромная генетическая разнородность. Четвериков это выразил так: «Вид, как губка, впитывает в себя гетерозиготные геновариации, сам оставаясь при этом все время внешне (фенотипически) однородным». Для жизни популяций эта особенность может иметь два разных следствия. В огромном большинстве случаев при изменении условий среды вид может реализовать свой «мобилизационный резерв» генетической изменчивости не только за счет новых наследственных изменений у каждой особи, но и благодаря «генетическому капиталу», доставшемуся от предков. Благодаря такому механизму наследования популяция приобретает пластичность, без чего невозможно обеспечить устойчивость приспособлений в меняющихся условиях среды. Однако изредка возможен и другой исход: редкие скрытые вредные мутации иногда могут встретиться у потомства совершенно здоровых родителей, приводя к появлению особей с наследственными заболеваниями. И это – тоже закономерное, неистребимое биологическое явление, своего рода жестокая плата популяции за поддержание своей наследственной неоднородности.
С. С. Четверикову популяционная генетика обязана еще одним открытием, которое было изложено в маленькой, всего на 4 страницы, заметке «Волны жизни», опубликованной в 1905 году на страницах «Дневника Зоологического отделения Императорского общества любителей естествознания и этнографии» в Петербурге. Он обратил внимание, что поскольку любая природная популяция имеет конечную, ограниченную численность особей, это неизбежно приведет к чисто случайным статистическим процессам в распространении мутаций. При этом популяции всех видов постоянно меняют численность (численность грызунов в лесу может от года к году изменяться в сотни, а многих видов насекомых – в десятки тысяч раз), из-за чего в разные годы распространение мутаций в популяциях может идти совершенно по-разному. От громадной популяции птиц, насекомых, зайцев и других животных в трудный для переживания год может остаться всего несколько особей, причем иногда совершенно нетипичных для бывшей популяции. Но именно они дадут потомство и передадут ему свой генофонд, так что новая популяция по составу генетического материала будет совершенно иной, чем прежняя. В этом проявляется генетический «эффект основателя» популяции.
Постоянно изменяется и геном в популяциях человека. К. Альстрем на материале в южной Швеции показал, что в популяции человека передается следующему поколению далеко не весь имеющийся генофонд, а лишь избранная, а то и случайно «выхваченная» часть. Так, 20% поколения здесь вовсе не оставили потомков, зато 25% родителей, которые имели трех и более детей, дали 55% численности следующего поколения.
Таким образом, в большой полиморфной панмиктической популяции, состоящей из одинаково жизнеспособных и одинаково плодовитых особей, различные гомозиготные и гетерозиготные генотипы быстро достигают некоторых равновесных частот, зависящих от существующих в популяции частот аллелей. Частоты генотипов, достигнув равновесия Харди-Вайнберга, остаются затем постоянными на протяжении всех последующих поколений при случайном скрещивании. Частоты аллелей в большой полиморфной популяции, состоящей из одинаково жизнеспособных и одинаково плодовитых особей, обычно остаются постоянными из поколения в поколение. Это постоянство не зависит от случайного скрещивания. Поэтому применительно к частотам аллелей закон Харди — Вайнберга носит более общий характер, чем применительно к частотам генотипов, и в этом своем аспекте он имеет более важное значение для эволюционной теории.
Необходимо проверять особые условия, необходимые для действия закона Харди — Вайнберга. Эти условия исключают любые факторы, воздействующие на частоту генов, кроме самого процесса воспроизведения генов. До тех пор пока уровень изменчивости в генофонде определяется воспроизведением генов как таковым, этот уровень остается постоянным и неизменным в ряду последовательных поколений. Таким образом, закон Харди-Вайнберга состоит из следующих двух утверждений. Частоты гамет (аллелей) не меняются от поколения к поколению. Равновесные частоты генотипов достигаются за одно поколение. В популяции поддерживается соотношение между гомозиготными и гетерозиготными организмами. [1]
Заключение
Английский математик Харди и немецкий врач Вайнберг примерно одновременно в 1908 году доказали основополагающую теорему популяционной генетики, которая объясняет, почему от поколения к поколению не возрастает частота встречаемости доминантных генов. Закономерность наследования признаков в популяциях раздельнополых животных и перекрестноопыляемых растений, получившая название закона Харди-Вайнберга, отражает зависимость между частотами аллелей и генотипов в популяциях. Данный закон объясняет, каким образом в популяции сохраняется генетическое равновесие, то есть число особей с доминантными и рецессивными признаками остается на определенном уровне.
Согласно этому закону, частоты доминантных и рецессивных аллелей в популяции будут оставаться постоянными из поколения в поколение при наличии определенных условий: высокой численности особей в популяции; свободном их скрещивании; отсутствии отбора и миграции особей, а также одинаковой численности особей с разными генотипами.
Нарушение хотя бы одного из этих условий ведет к вытеснению одного аллеля (например, A) другим (а). Под действием естественного отбора, популяционных волн и других факторов эволюции особи с доминантным аллелем А будут вытеснять особи с рецессивным аллелем а.
В популяции может измениться соотношение особей с разными генотипами. Предположим, что генетический состав популяции был таким: 20% АА, 50% Аа, 30% аа. Под воздействием факторов эволюции он может оказаться следующим: 40% АА, 50% Аа, 10% аа. Используя закон Харди-Вайнберга, можно вычислить частоту встречаемости любого доминантного и рецессивного гена в популяции, а также любого генотипа.
На первый взгляд может показаться, что особи с доминантным фенотипом должны встречаться чаще, чем с рецессивным. Однако соотношение 3:1 соблюдается лишь в потомстве двух особей, гетерозиготных по одним и тем же двум аллелям. При других типах скрещивания в потомстве происходит иное расщепление признаков, и такие скрещивания также влияют на частоты генотипов в популяции. Законы Менделя ничего не говорят нам о частотах генотипов в популяциях. Именно об этих частотах идет речь в законе Харди – Вайнберга.
Основное утверждение закона Харди – Вайнберга состоит в том, что в отсутствие элементарных эволюционных процессов, а именно мутаций, отбора, миграции и дрейфа генов, частоты генов остаются неизменными из поколения в поколение. Этот закон утверждает также: если скрещивание случайно, то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями.
Из закона Харди – Вайнберга вытекает следующий вывод: если частоты аллелей у самцов и самок исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигаются за одно поколение. Если частоты аллелей у двух полов исходно различны, то для аутосомных локусов они становятся одинаковыми в следующем поколении, поскольку и самцы, и самки получают половину своих генов от отца и половину – от матери. Таким образом, равновесные частоты генотипов достигаются в этом случае за два поколения. Однако в случае сцепленных с полом локусов равновесные частоты достигаются лишь постепенно.
Список использованных источников:
1. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т.3. – М.: Мир, 1987.
2. География и мониторинг биоразнообразия. Коллектив авторов. М.: Издательство Научного и учебно-методического центра, 2002. – 432 с.
3. Грант Верн. Эволюционный процесс. – М.: Мир, 1989.
4. Равич-Щербо И.В., Марютина Т.М., Григоренко Е.Л. Психогенетика: Уч. Для вузов. – М.: Аспект Пресс, 2000. – 447 с.
5. Словарь терминов по генетике. http://www.glossary.ru
6. Фогель Ф., Мотульски А. Генетика человека: В 3-х т. Т.1: Пер. с англ. – М.: Мир, 1989. – 312 с.
Похожие работы
... . Совокупность сцепленных генов одной хромосомы, контролирующих аллогруппу, называют гаплотипом. Значение: 1) изучение причин и динамики генотипической изменчивости, составляющей основу эволюционной генетики; 2) уточнение происхождения отдельных животных; 3) определения моно- и дизиготных двоен; 4) построение генетических карт хромосом; 5) использование биохимических систем в качестве генетических ...

... поколений. Естественно, особенно они заметны, если популяция находится в изоляции, т.е. отсутствует миграция генов извне. Известны сообщества такого рода в человеческом обществе. Часть 2 Математические модели нейронных систем Изучение нейронных систем -одно из самых романтических направлений научных исследований, поскольку нейронные системы присущи как человеку, так и животным. Самая ...

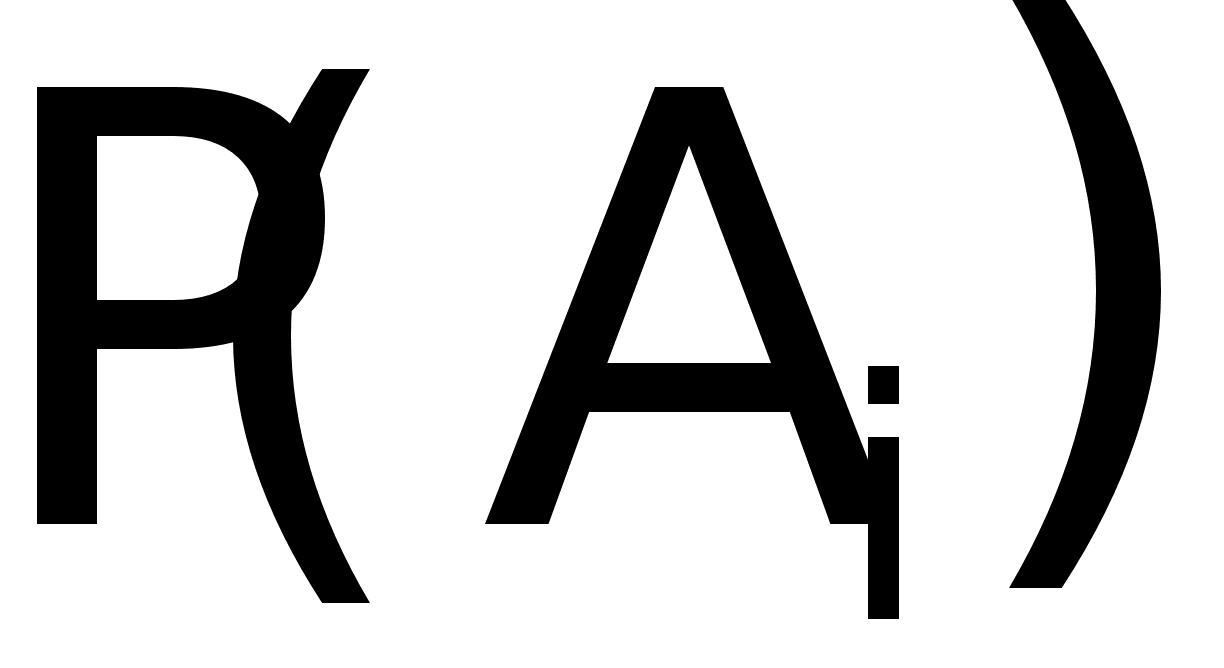
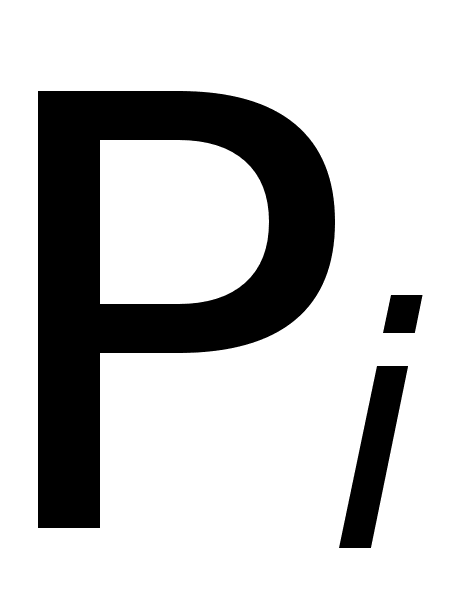
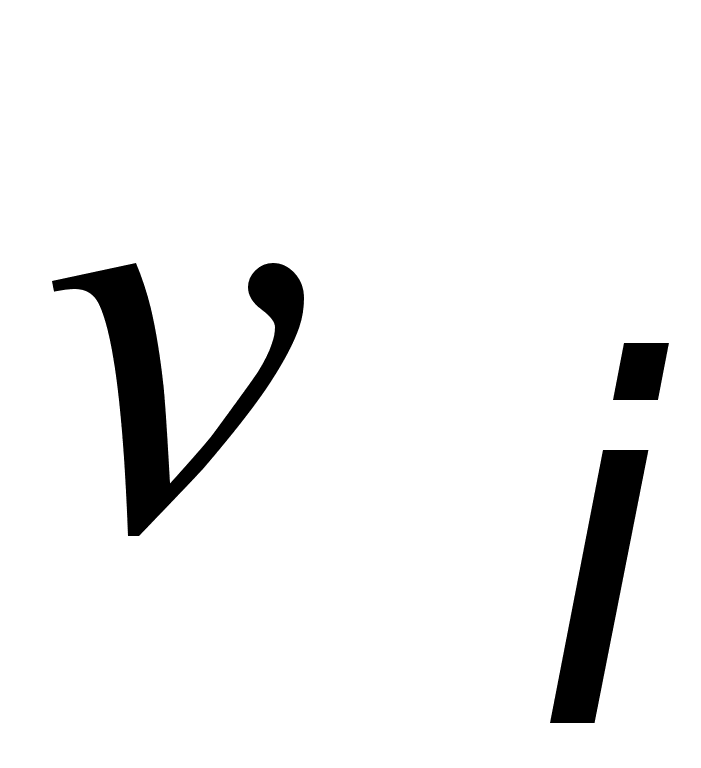
... показывают: Таким образом, . Совершенно аналогично: , , . В равновесных состояниях частоты гамет являются произведениями частот соответствующих генов. Верно и обратное утверждение. Часть 2 Математические модели нейронных систем Изучение нейронных систем -одно из самых романтических направлений научных исследований, поскольку нейронные системы присущи как человеку, так и животным. Самая ...
... аллеля равна 0.99, а частота рецессивного аллеля альбинизма – 0.01. Общая частота аллелей в популяции составляет 100%, или 1.0, поэтому Частота доминантного аллеля + Частота рецессивного аллеля =1 0.99 + 0.01 =1 Как это принято в классической генетике, аллели можно обозначить буквами, например, доминантный аллель (нормальная ...
0 комментариев