Навигация
Математические модели естествознания
143087
знаков
0
таблиц
437
изображений
Курс лекций
Математические модели в естествознании
1996 г. 1
Часть 1
Основы математической генетики 2
Законы Менделя 4
Закон Харди- Вайнберга 8
Курс лекций
Математические модели в естествознании
1996 г.
Работа над любой математической моделью начинается со сбора и анализа фактического материала. Определяются цели моделирования. Выделяются главные черты изучаемого объекта или явления. Вводятся формализованные характеристики. Принимаются правила работы с ними. В результате возникает математический объект, который и называется математической моделью. Разрабатываются методы математического анализа модели, которыми она исследуется. Полученные результаты математического моделирования интерпретируются в рамках исходного фактического материала, что позволяет оценить степень адекватности модели. Результаты моделирования не должны противоречить выделенным ранее ключевым экспериментальным фактам. Одновременно, модель не может объяснить все стороны изучаемого объекта или явления.
Хорошая модель, кроме объяснения известных, должна давать возможность предсказывать новые свойства. Математическое моделирование широко используется там, где экспериментальные исследования трудоемки и дорогостоящи, или вообще невозможны (например, в изучении социальных явлений).
Кроме задачи о прогнозе, математическое моделирование помогает классифицировать и систематизировать фактический материал, увидеть существующие связи в мозаике фактов. Это вытекает из того, что модель является специфическим -ярким и выразительным языком, предназначенным для описания для описания изучаемого объекта или явления.
Мир математических моделей разнообразен. Существуют различные схемы их классификации. Однако, каждая модель конкретна и предназначена для описания достаточно узкого круга объектов и явлений. Это накладывает определенный отпечаток на предлагаемый лекционный курс. Он включает в себя модели. относящиеся к различным областям естествознания. Модели сильно отличаются друг от друга не только предметными областями, но математической терминологией, а также математическими методами их исследования.
Предпочтение отдается более простым моделям. Отметим, что "простота" (иногда в ущерб точности)-один из принципов, о котором всегда нужно помнить при разработке математической модели.
Часть 1Основы математической генетики
В 1865 г. чешский монах Грегор Мендель опубликовал работу о результатах скрещивания разновидностей гороха. В своих опытах Г. Мендель изучал закономерности наследования семи пар альтернативных признаков. В одном из опытов перекрестно скрещивались растения с гладкими и сморщенными семенами. В результате такого скрещивания в первом поколении все растения имели гладкие семена. Проявляющиеся признаки Г.Мендель назвал доминантными, а не проявляющиеся - рецессивными. Растениям, полученным в первом поколении, была предоставлена возможность самоопыляться. Во втором поколении появились как гладкие, так и сморщенные горошины. При подсчете выяснилось, что 5 474 горошины были гладкими, а 1 850 - морщинистыми. Доля гладких горошин оказалась близкой к 3/4, а морщинистых - к 1/4. Отношение близко к 3:1. Во всех других опытах для каждой пары альтернативных признаков (например, цвет горошин) доминантный признак обнаруживался примерно втрое чаще рецессивного. Для объяснения результатов опытов Г.Мендель предположил, что внешние признаки определяются некоторыми внутренними факторами (генами), которые могут находиться в одной из двух альтернативных форм (теперь они называются аллелями). Были сделаны два допущения.
1. Два фактора, определяющие данный признак, в течении жизни организма сосуществуют независимо друг от друга, не сливаясь и не растворяясь один в другом. Они расщепляются при формировании половых клеток, которые возникают парами. Одна из половых клеток несет в себе один, а другая - оставшийся фактор.
2. Гены, определяющие различные признаки, наследуются независимо.
Работа Г.Менделя была не понята и забыта его современниками. В 1900 г. результаты были открыты вновь. Началось развитие новой науки -генетики. Согласно современным представлениям, носителями генов являются нитевидные тела -хромосомы, которые располагаются в ядре клетки. Организмы, относящиеся к различным биологическим видам имеют разную структуру и число хромосом. У человека каждая клетка содержит 46 хромосом, у гороха -14. Число хромосом четно. Каждая хромосома в клетке присутствует в двух экземплярах, отличающихся, быть может только аллелями (вариантами, или формами) генов. Такие пары хромосом называются гомологичными. Место, занимаемое аллелью гена в хромосоме, называется локусом. Можно представить хромосому как прямолинейный отрезок, а локусы как его последовательные участки.
Половые клетки -гаметы возникают в результате процесса, который называется мейозом. Гомологичные хромосомы расходятся в разные концы клетки, и клетка делится пополам. Гаметы содержат половинный набор хромосом (у человека 23). Зародышевая клетка -зигота образуется путем слияния мужской и женской гамет и содержит полный набор хромосом. Из зиготы путем обычного деления - митоза образуется новый организм. При митозе каждая хромосома создает свою точную копию. Оригиналы и копии расходятся в разные клетки. Набор генов каждой клетки называется генотипом организма. Описанная схема упрощена. но в ней отражаются ключевые моменты, которые используются при разработке математической модели.
Фенотипом называется совокупность всех внешних признаков организма. Фундаментальный принцип генетики - при неизменных внешних условиях фенотип организма определяется его генотипом. Некоторые признаки организма определяются не всем не всем генотипом, а только его частью (в экспериментах Г.Менделя - двумя аллелями одного гена). Пол человека определяется двумя половыми хромосомами X и Y. Женская зигота содержит две хромосомы X, мужская -хромосомы X и Y.
Мать передает своему ребенку одну из хромосом X. Отец передает дочери хромосому X, а сыну - хромосому Y. Таким образом, пол ребенка зависит только от отца.
Совокупность генотипов, у которых часть генных наборов одинакова, называется популяцией (по данным наборам). Численность популяции считается достаточно большой (бесконечной). Описывать популяцию будем набором частот генотипов в данном поколении. Будем считать, что одно поколение сменяет другое, т.е. поколения не перекрываются.
Законы МенделяРассмотрим рассуждения Г.Менделя, используя описанные выше термины. За форму семян гороха отвечает двухаллельный ген. Его доминантную аллель (фенотип -гладкие семена) обозначим как , а рецессивную как a (сморщенные семена). Генотип определяется парой аллелей. Возможны три генотипа: AA, Aa, aa. Генотип Aa называется гетерозиготным, а генотипы AA, aa -гомозиготными. Поскольку аллель A является доминантным, то растения первых двух генотипов будут иметь гладкие семена, а третьего -морщинистые.
На первом этапе своего опыта Г.Мендель брал гомозиготные растения AA, aa. Первый тип давал гамета, имеющие аллель A, второму соответствовали гаметы с аллелью a.
Скрещивание гомозиготных растений AA и aa (слияние гамет A и a) дает гетерозиготное растение Aa. Последнее дает гаметы, несущие аллели A и a. Какую из двух аллелей получит конкретная гамета -дело случая и вероятность каждого из событий 1/2.
На втором этапе опыта Г.Менделя скрещивались гетерозиготные растения. Генотип AA у потомка имеет место, если каждый из родителей передал гамету с аллелем A. События независимые. Вероятность P(AA) потомка с генотипом AA равна 1/4. Аналогично, вероятность P(aa) появления потомка aa также 1/4. Вероятность появления потомка с генотипом Aa можно вычислить по дополнению: P(Aa)=1-1/4-1/4=1/2. Вероятность того, что растение -потомок двух гетерозиготных родителей будет иметь гладкие семена: P(AA)+P(Aa)=1/4+1/2=3/4. Морщинистые семена будут наблюдаться с вероятностью P(aa)=1/4.
Такова математическая модель, объясняющая опыты Г.Менделя. Обсудим вопрос об ее адекватности. Напомним, что в опытах соответствующие частоты наблюдались приближенно. Это задача о проверке статистической гипотезы. Для проверки можно использовать критерий Пирсона.
Предположим,
что мы наблюдаем
серию из n независимых
испытаний.
Каждое из них
может завершиться
одним из m исходов
![]() .
Вероятности
исходов
.
Вероятности
исходов ![]() не меняются
от испытания
к испытанию.
Подлежащая
проверке нуль
-гипотеза состоит
в том, что эти
вероятности
равны некоторым
заранее заданным
числам:
не меняются
от испытания
к испытанию.
Подлежащая
проверке нуль
-гипотеза состоит
в том, что эти
вероятности
равны некоторым
заранее заданным
числам: ![]() .
Относительно
числа n предполагается.
что оно достаточно
велико. Пусть
.
Относительно
числа n предполагается.
что оно достаточно
велико. Пусть
![]() -полученные
в результате
опыта эмпирические
частоты наступления
исхода
-полученные
в результате
опыта эмпирические
частоты наступления
исхода ![]() .
Составляется
сумма:
.
Составляется
сумма:
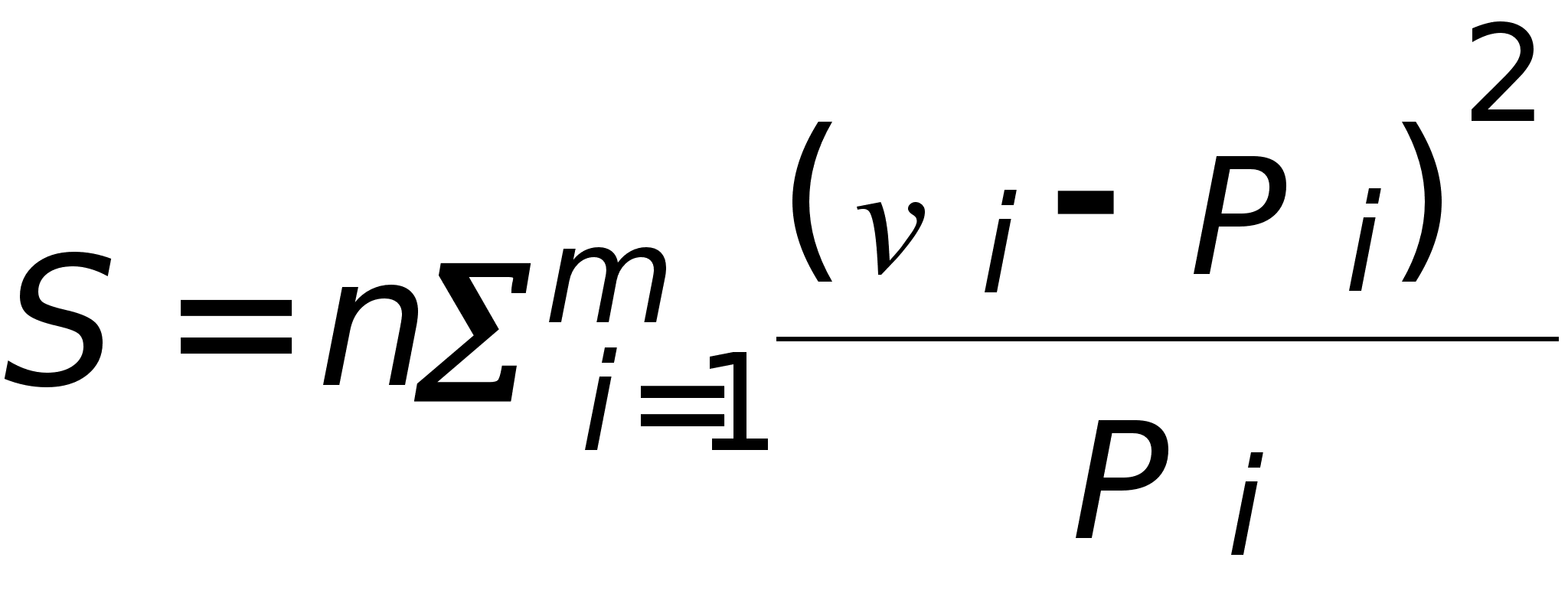 ,
,
которая
часто называется
суммой Пирсона.
Оказывается,
что с ростом
n распределение
статистики
S стремится к
предельному
распределению
![]() с m-1 степенями
свободы, не
зависящему
ни от n, ни от чисел
с m-1 степенями
свободы, не
зависящему
ни от n, ни от чисел ![]() .
Для любого e>0
можно указать
практическую
границу
.
Для любого e>0
можно указать
практическую
границу ![]() такую, что
такую, что
![]() .
.
Другими
словами, неравенство
![]() >
>![]() практически
невозможно.
Число e
называется
уровнем значимости.
практически
невозможно.
Число e
называется
уровнем значимости.
Зададимся
уровнем значимости
e=0.05, который
очень популярен
в биологии. В
рассматриваемом
случае число
степеней свободы
m-1=1. Распределение
суммы Пирсона
для столь большого
числа, как n=7 324
практически
не отличается
от ![]() распределения
с одной степенью
свободы. По
таблицам определяем
распределения
с одной степенью
свободы. По
таблицам определяем
![]() »3.84.
Вычислим сумму
Пирсона:
»3.84.
Вычислим сумму
Пирсона:
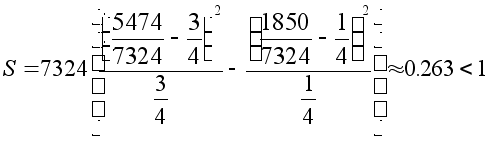
Так
как сумма Пирсона
меньше ![]() ,
то нуль- гипотеза
хорошо согласуется
с результатами
эксперимента.
Получаем веские
основания
считать, что
закон Менделя
справедлив.
,
то нуль- гипотеза
хорошо согласуется
с результатами
эксперимента.
Получаем веские
основания
считать, что
закон Менделя
справедлив.
В генетике развит некий формализм, позволяющий быстро выводить закономерности. Аллельные пары представляют в виде:
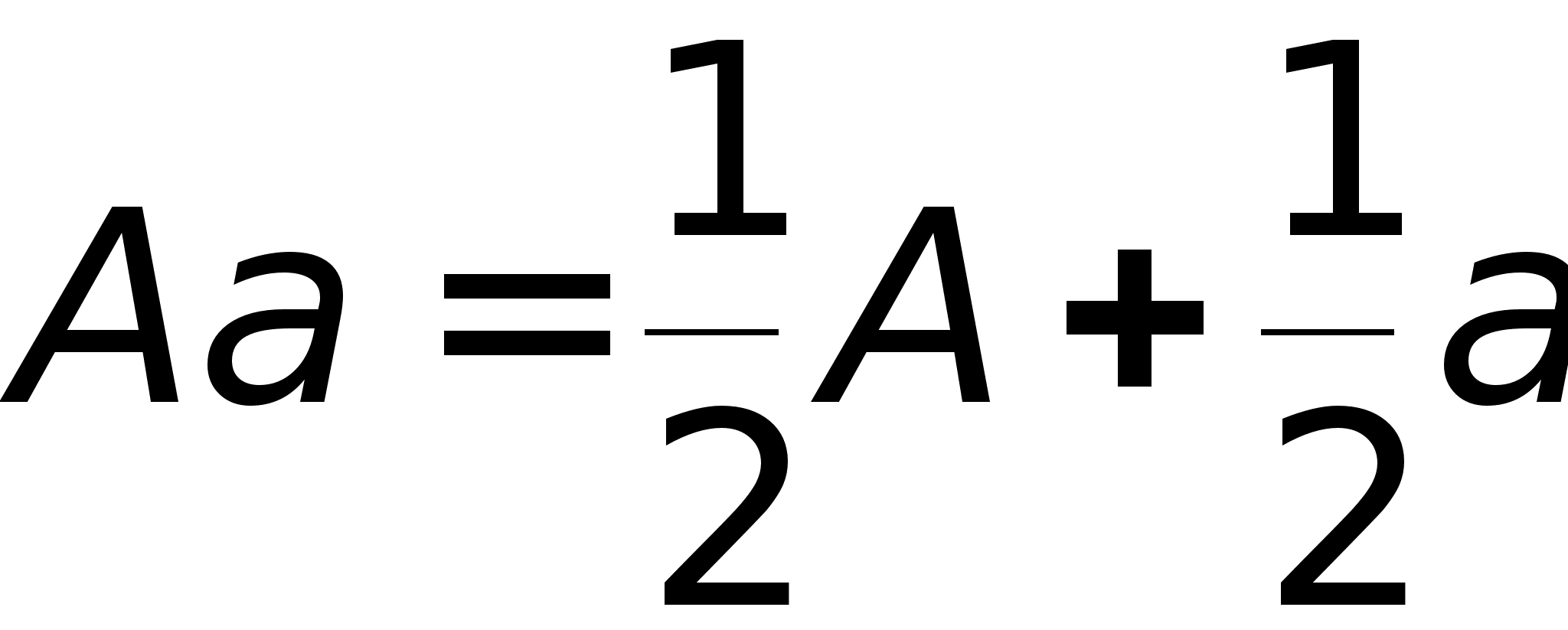 ,
,
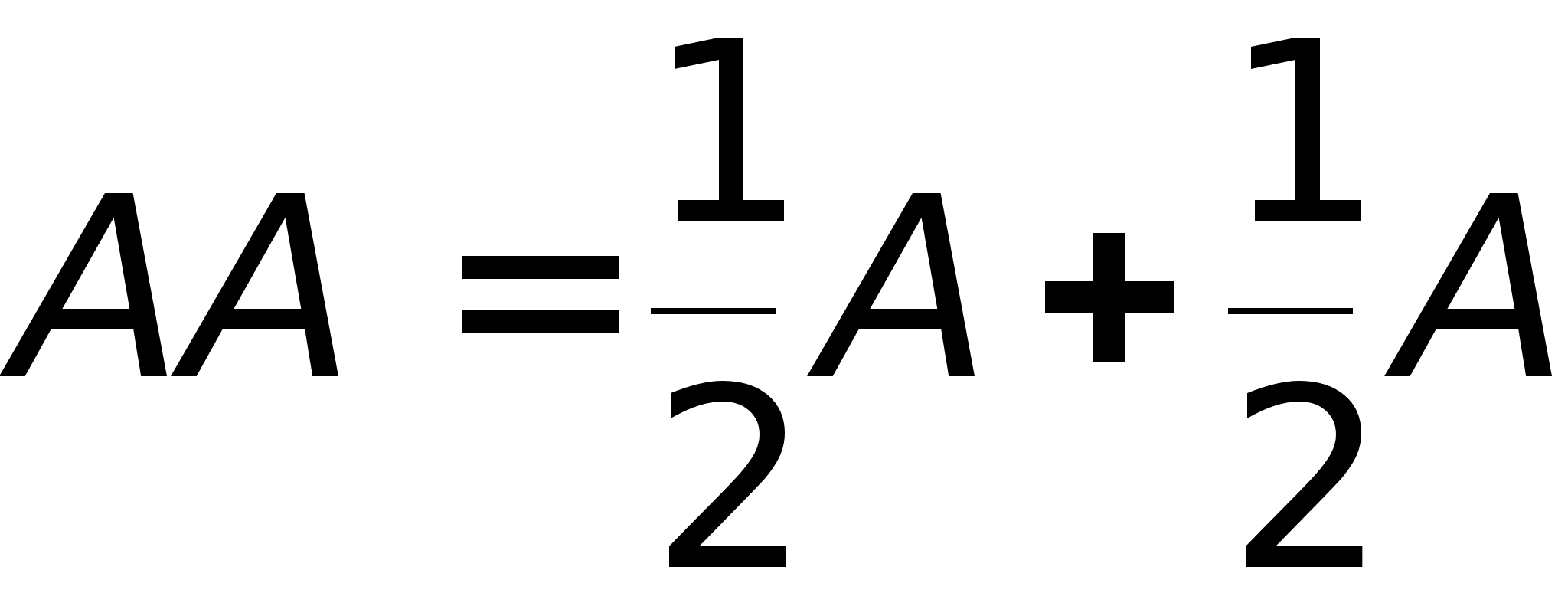 ,
,
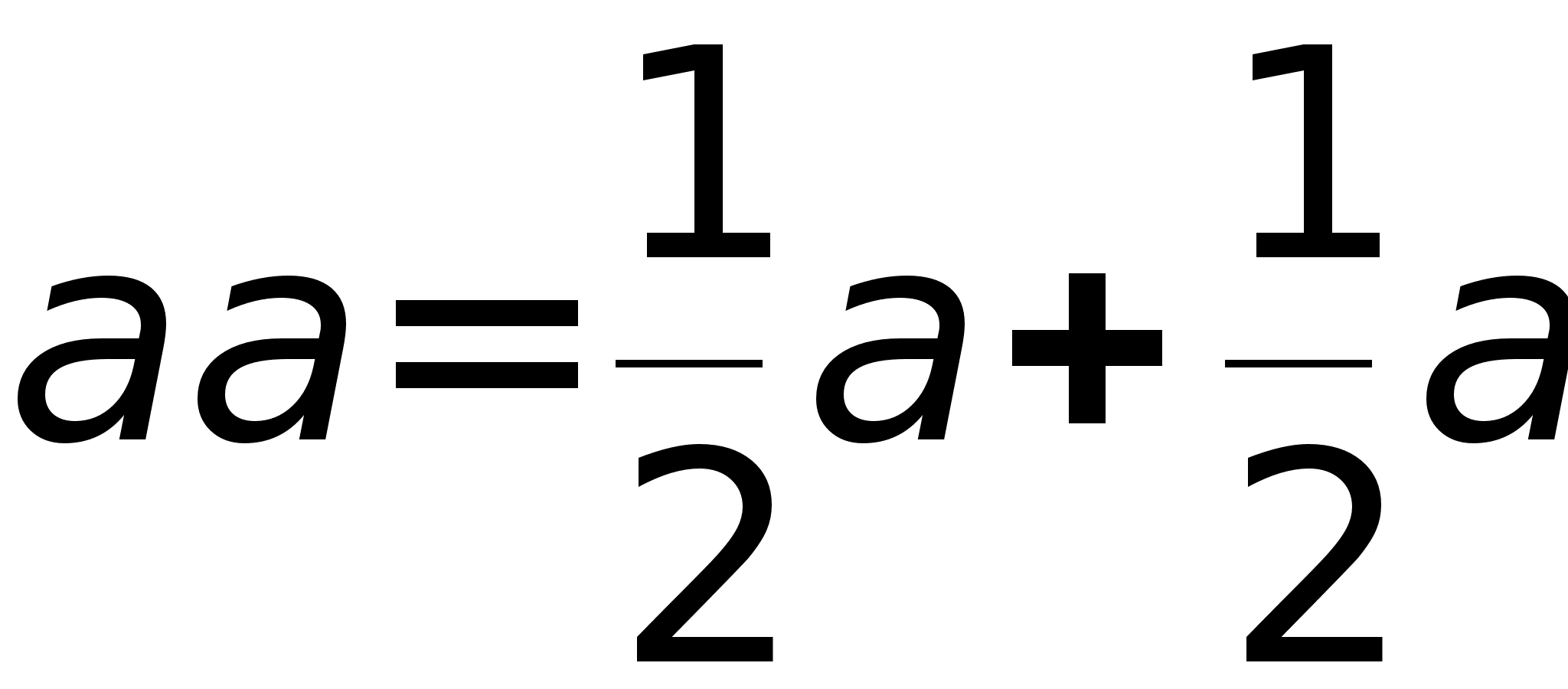 ,
,
Скрещивание генотипов обозначается знаком 'ґ' -умножения. Скобки в формулах раскрываются по привычным правилам и знаки умножения опускаются. Скрещивание гетерозиготных растений описывается формулой:
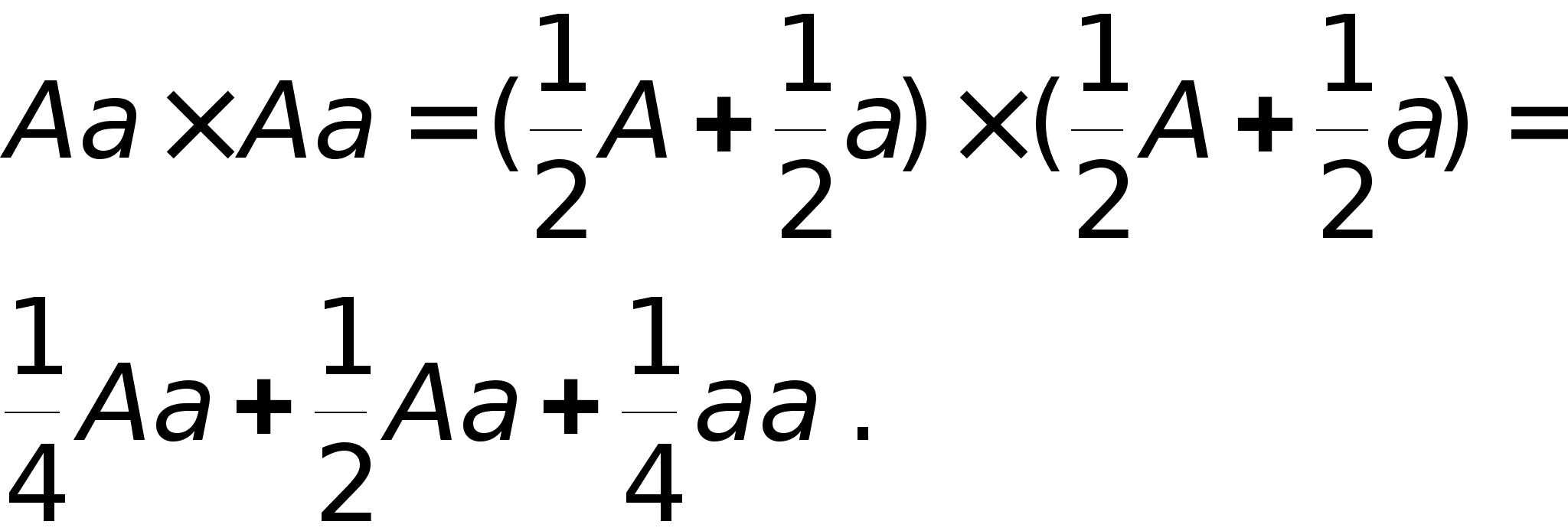
Полученная формула утверждает, что генотипы AA и aa возникают при скрещивании с вероятностью 1/4, а генотип Aa с вероятностью 1/2. Так как генотипы AA и Aa обладают гладкими семенами, то 3/4 потомства имеют гладкие семена, а 1/4 - морщинистые семена (генотип aa).
Решим простую задачу о скрещивании генотипов Aa и aa:
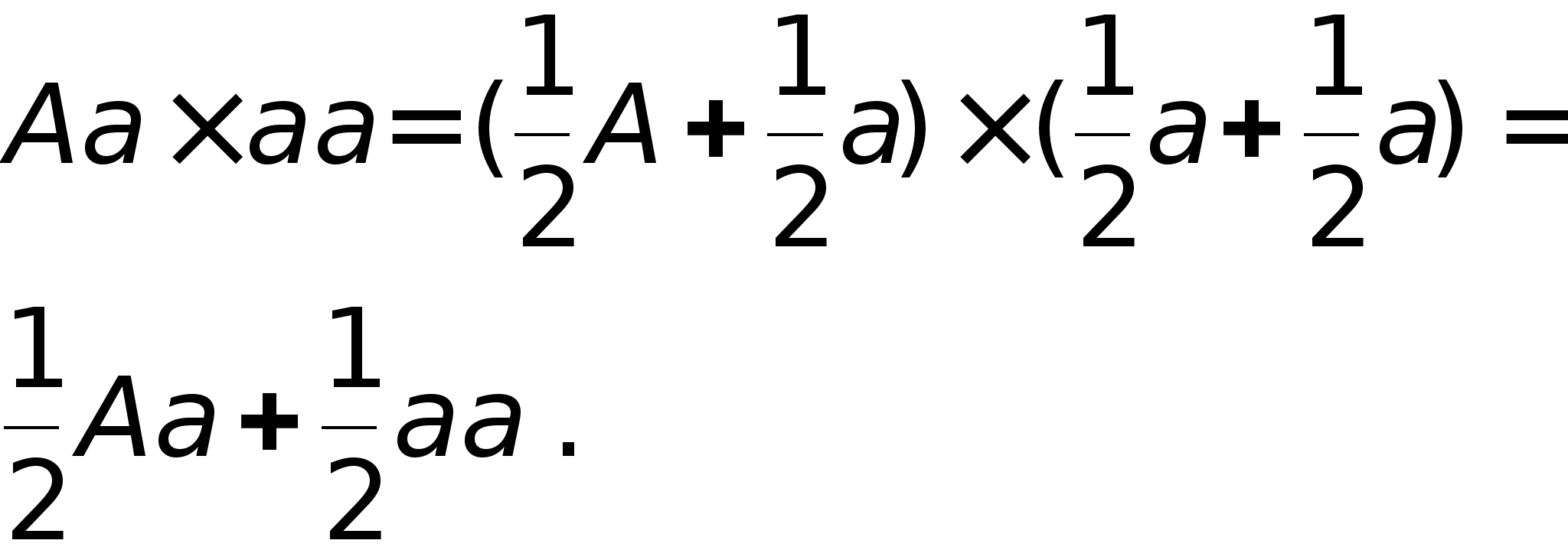
Таким образом, половина генотипов будет гетерозиготными, а половина гомозиготными.
Большинство признаков генотипа контролируется более чем двумя аллелями. Такие аллели называются множественными. Такие аллели в любом непарном сочетании могут находиться в любой клетке, так как только две аллели одного гена могут одновременно присутствовать в генотипе. Такие генотипы называются диплоидными. Полиаллельными являются гены, контролирующие группы крови. Группа крови человека зависит от присутствия либо отсутствия в эритроцитах специфических белков (A и B). Существуют четыре группы крови: Группа крови A с генотипами AA и AO (группа крови содержит белок A), группа крови B с генотипами BB и BO (содержит белок B), Группа крови AB (содержит оба белка), группа крови OO (отсутствие белков A и B). Таким образом, группа крови контролируется тремя аллелями A, B, O одного гена. Аллели A и B -доминанты по отношению к O. В присутствия аллелей A и B доминантность отсутствует. Таким образом группы крови определяются шестью генотипами AA, AO, AB, BB, BO, OO.
Закон Харди- Вайнберга
В законе Харди -Вайнберга речь идет о частотах генотипов в популяциях. Этот закон сформулировали в 1908 г. независимо друг от друга английский математик Дж.Харди и австрийский врач В.Вайнберг. Рассматривалась следующая задача. Известны частоты генотипов в двухаллельной популяции в нулевом поколении. Требуется проследить изменение частот от поколения к поколению.
Двухаллельная популяция состоит из генотипов: AA, Aa, aa. Их частоты в нулевом (начальном) поколении обозначим через u(0), 2v(0), w(0). Естественно, что u(0)+2v(0)+w(0)=1. Скрещивание предполагается случайным. Удобно следить за эволюцией частот с помощью следующей схемы.
Нулевое поколение
Генотипы Частоты генотипов
AA u(0)
Aa 2v(0)
aa w(0)
Гаметы Частоты гамет
A p(0)=u(0)+v(0)
a q(0)= v(0)+w(0)
(менделевское формирование гамет)
Первое поколение
Генотипы Частоты генотипов![]()
AA ![]()
![]()
Aa ![]()
aa ![]()
Гаметы Частоты гамет
A
![]()
a ![]()
Таким образом, частоты гамет не меняются от поколения к поколению. Дальнейшее скрещивание не меняет и частоту зигот.
Второе поколение
Генотипы Частоты генотипов![]()
AA ![]()
![]()
Aa ![]()
aa ![]()
Частоты зигот устанавливаются в первом поколении и больше не меняются.
Закон Харди-Вайнберга состоит из следующих двух утверждений.
Похожие работы
... моделирование широко используется там, где экспериментальные исследования трудоемки и дорогостоящи, или вообще невозможны (например, в изучении социальных явлений). Кроме задачи о прогнозе, математическое моделирование помогает классифицировать и систематизировать фактический материал, увидеть существующие связи в мозаике фактов. Это вытекает из того, что модель является специфическим -ярким и ...

... поколений. Естественно, особенно они заметны, если популяция находится в изоляции, т.е. отсутствует миграция генов извне. Известны сообщества такого рода в человеческом обществе. Часть 2 Математические модели нейронных систем Изучение нейронных систем -одно из самых романтических направлений научных исследований, поскольку нейронные системы присущи как человеку, так и животным. Самая ...
... если мы увидели одного тигра, то не кишат ли все джунгли тиграми, и кто знает, где притаился следующий?». Примером одной из моделей, обсуждаемых как минимум в течение более чем двух столетий, является модель демографического взрыва: в конце 18 века Мальтус впервые поднял вопрос о росте численности человечества по экспоненте. Экспоненциальный рост описывается уравнением (где N – численность ...
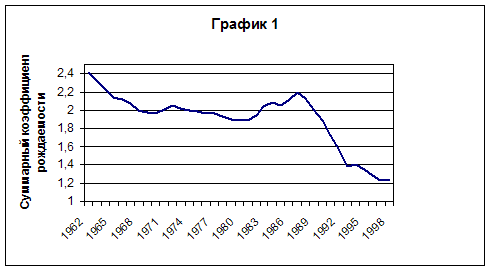
... и применяет математические методы для изучения взаимосвязей демографических явлений и процессов, моделирования и прогнозирования. В числе демографических моделей - вероятностные таблицы смертности, брачности, рождаемости, модели стационарного и стабильного населения, имитационные модели демографических процессов и т. п. - историческая демография; которая изучает состояние и динамику ...
0 комментариев