Нейропептиды и их обмен
Физико-химические свойства, субстратная специфичность и биологическая роль карбоксипептидазо-В-подобных ферментов
ФМСФ-ингибируемая карбоксипептидаза
Половые стероидные гормоны
Прогестерон
Методы исследования
Метод определения активности карбоксипептидазы Н
Распределение активности ФМСФ-ингибируемой карбоксипептидазы в в гипоталамо-гипофизарно-надпочечниково-гонадной системе самцов и самок мышей
Активность карбоксипетидазы Н и ФМСФ-ингибируемой карбоксипептидазы при введении прогестерона в тканях самок мышей
Активность основных карбоксипептидаз у самцов мышей при введении тестостерона и прогестерона
Навигация
Половые стероидные гормоны
Активность основных карбоксипептидаз в тканях мышей при введении тестостерона и прогестерона
168432
знака
5
таблиц
0
изображений
1.3. Половые стероидные гормоны
Гормоны служат важными регуляторами различных физиологических процессов, а также поведения животных и человека. Действие этих биологически активных веществ распространяется на все органы и системы организма. Половые железы продуцируют довольно много стероидов, но лишь незначительная их часть обладает гормональной активностью. Образование этих гормонов строго регулируется с помощью петли обратной связи, включающей в себя гипофиз и гипоталамус. Необходимо отметить, что образование половых стероидов не является прерогативой гонад. Определенный вклад в этот процесс вносят и надпочечники. Именно поэтому часто гипоталамус (источник рилизинг-факторов), гипофиз (орган, в котором синтезируются ЛГ, ФСГ), надпочечники и половые железы (основные источники половых стероидов) объединяют в одну систему – гипоталамо-гипофизарно-надпочечниково-гонадную (ГГНГС), которая участвует в регулировании важных биологических функций организма.
Одними из половых стероидных гормонов, влияющих на уровнь регуляторных пептидов, являются тестостерон и прогестерон [32, 164, 175, 245, 266].
1.3.1. Тестостерон
Тестостерон является основным мужским половым гормоном (в норме на его долю приходится около 90% от всех секретируемых андрогенов [27]). 90% этого гормона продуцируется семенниками, 5% – корой надпочечников и 5% образуется в результате метаболизма других стероидных гормонов в неэндокринных органах [27].
Тестостерон, как и другие андрогены, относится к группе С19-стероидов (андростановая группа). Его синтез в клетках Лейдига регулируется лютеинизирующим гормоном (ЛГ) передней доли гипофиза.
Скорость-лимитирующим этапом в синтезе тестостерона является отщепление боковой цепи холестерина. Основной путь образования прогестероновый или Δ4-путь: прегненолон à прогестерон à 17a-гидроксипрогестерон à андростендион à тестостерон [24], причем превращение холестерина в прегненолон при участии цитохрома-Р450, отщепляющего боковую цепь (Р-450обц), в надпочечниках, яичниках и семенниках происходит идентично [32].
В крови большинства млекопитающих (в том числе и человека) циркулирующий гормон на 97-99% связан с белками плазмы, преимущественно с b-глобулином – секс-гормон-связывающим глобулином или тестостерон-эстроген-связывающим глобулином, а также с альбумином [24, 27, 32]. В семенниках тестостерон специфически связан с андрогенсвязывающим белком, синтезируемым под влиянием фолликулостимулирующего гормона (ФСГ) передней доли гипофиза в клетках Сертоли. Благодаря андрогенсвязывающему белку, достигается большая, по сравнению с ситемной кровью, концентрация тестостерона, которая необходима для созревания и дифференцировки половых клеток [24, 32]. Секс-гормон-связывающий глобулин связывает тестостерон с относительно высоким сродством и ограниченной емкостью [32]. Следует отметить, что связывание тестостерона с СГСГ происходит с большим сродством, чем других половых стероидных гормонов, например эстрадиола. Поэтому колебания количества данного белка в плазме крови существенно отражаются на концентрации свободного тестостерона, как раз и обладающего биологической активностью [32, 205].
Период полураспада тестостерона в плазме крови колеблется от 40 до 100 мин [42, 192]. Метаболическая деградация тестостерона протекает двумя основными путями. Первый – наиболее эффективный – происходит, главным образом, в печени. Он заключается в окислении атома углерода в 17-положении. В результате образуются 17-кетостероиды (17-оксостероиды), полностью лишенные или обладающие более слабой активностью, чем исходное соединение. Эти метаболиты в значительной части конъюгируют с сульфатом, с образованием водорастворимых соединений. В этом состоянии они выводяться через почки [24, 27, 32]. Часть 17-кетостероидов эксекретируется с желчью в кишечник [27].
При реализации второго пути инактивации тестостерона происходит восстановление двойной связи кольца А и 3-кетогруппы. При этом образуется активный метаболит – дигидротестостерон (ДГТ). Данный процесс катализируется НАДФН-зависисмой 5a-редуктазой и осуществляется непосредственно в тканях-мишенях [32]. Необходимо отметить, что ДГТ обладает большей по сравнению с тестостероном андрогенной активностью, большим сродством с СГСГ, и является тем соединением, посредством которого тестостерон осуществляет свое воздествие на ткани-мишени [32, 39].
Небольшая часть (1-5%) тестостерона подвергается ароматизации и превращается в эстрадиол. Данный процесс особенно важен для мозга, где эти гормоны участвуют в формировании полового поведения [24, 32, 39, 60, 166, 233].
У некоторых видов животных 5b-производные тестостерона стимулируют образование эритроцитов [39], но в основном 5b-ДГТ является неактивным метаболитом [111].
Основной физиологический эффект тестостерона – анаболический, то есть гормон обладает способностью стимулировать или ингибировать различные ферменты, участвующие в синтезе белка [24]. Этот гормон через андрогеновые рецепторы принимает участие в контроле развития жировой ткани [97], ингибирует экспрессию липопротеин-липазы и, при наличии гормона роста, заметно увеличивает липолиз через множественные взаимодействия по липолитическому каскаду [67]. Тестостерон увеличивает содержание b-серий ганглиозидов в почках крыс, но не влияет на содержание белков и сиалогликопротеидов [53]. Кроме того, данный гормон в высоких концентрациях обращает вызванный кастрацией эффект увеличения активности сиалотрансферазы I и сиалотрансферазы II в почках [53]. Способен значительно увеличивать продукцию секретируемого компонента в концевых клетках слезной железы крыс in vitro [261].
In vivo тестостерон снижает активность 11b-гидроксистероид дегидрогеназы [218]. Он повышает общий уровень гомоцистеина в плазме самок [129].
Интересно отметить влияние тестостерона, через ДГТ, на синтез ”простатического специфического антигена” (ПСА). Последний является гликопротеином, состоящим из одной полипептидной цепи (273 аминокислотных остатка) с пятью дисульфидными связями. Молекулярная масса с учетом микрогетерогенности по полисахариду составляет 30-33 кДа. ПСА в ткани предстательной железы и ее секрете обладает химотрипсинподобной активностью с каталитической триадой Gis41-Asp96-Ser189 и относится к сериновым протеазам [29].
Кроме того, ПСА обнаружен не только у мужчин, но и у женщин. В частности, в клетках рака молочной железы T47-D происходит синтез мРНК и белка ПСА. Однако это возможно лишь при стимуляции стериодными гормонами (прогестероном, ДГТ и в меньшей степени тестостероном и альдостероном). Эстрогены блокируют синтез ПСА [29].
Помимо этого тестостерон регулирует и множество других физиологических процессов через специфические рецепторы, число которых может повышаться либо понижаться [68, 78]. Этот гормон регулирует через ДГТ синтез андрогеновых рецепторов внутри клеток эндометрия [2], контролирует экспрессию 5a-редуктазы типа 2 в мозгу [201]. In vitro тестостерон снижает уровни мРНК рецепторов прогестерона, эстрогенов и андрогенов в клетках стромы эндометрия человека [162]. Этот гормон непосредственно (через андрогеновые рецепторы) подавляет экспрессию гена гонадотропин-рилизинг-фактора в гипоталамусе [63]. Он авторегулирует экспрессию мРНК собственных рецепторов в семенниках и стимулирует экспрессию андрогеновых рецепторов в клетках гранулезы растущих фолликулов [78, 278].
Интересно отметить тот факт, что при стрессе уровень тестостерона в плазме повышается. Причем секреция тестостерона стимулируется АКТГ [106].
В период половой зрелости у самок крыс тестостерон снижает связывание инсулина (посредством снижения числа поверхностных рецепторов в гепатоцитах) и еще более уменьшает его деградацию [167].
Ряд авторов отмечают тесную взаимосвязь между уровнями тестостерона, прогестерона и гормонами задней доли гипофиза (окситоцином, Arg8-вазопрессином и др.) [214, 266]. В частности тестостерон (в концентрации 0,01-0,1 мкг на мл), выступая в качестве предшественника 17b-эстрадиола, увеличивает in vitro секрецию окситоцина гранулезными клетками яичников. При более высоких концентрациях наблюдается обратный эффект – ингибирование секреции [274]. Тестостерон влияет на экспрессию пролактина в дорсальной и латеральной простате [213] и может выступать в качестве стимулятора секреции пролактин-подобного пептида клетками яичников [245]. Этот гормон в некоторой степени ингибирует высвобождение Arg8-вазопрессина из гипоталамических срезов in vitro [175]. Окситоцин, Arg8-вазопрессин и Arg8-вазотоцин участвуют в регуляции секреции тестостерона и прогестерона [214]. Кроме того, имеются данные о том, что уменьшение рецепторов окситоцина при старении в обонятельных луковицах и гипоталамическом вентромедиальном ядре связано со снижением уровня тестостерона в плазме [56].
Тестостерон (в отличие от прогестерона) не оказывает влияния на аккумуляцию кальцитонин-ген-зависимого пептида ни в ядрах тройничного нерва, ни в шейном отделе позвоночника [208]. С другой стороны, по мнению авторов [208], факт наличия кальцитонин-ген-зависимого пептида в гипоталамусе только самцов, свидетельствует о положительном эффекте тестостерона.
На клеточном уровне тестостерон способствует увеличению (а прогестерон – уменьшению) зоны гранулярных микротрубочек в клетках подчелюстной железы крыс [90], играет ведущую роль в регуляции транспорта Ca2+ в клетках миометрия [252], стимулирует продукцию цАМФ в клетках яичников свиньи [244].
Тестостерон и другие андрогены в виде 5a-редуцированных производных оказывают влияние на клетки ЦНС [200], вместе с прогестинами способствуя их дифференцировке на ранних этапах онтогенеза [193, 198]. При этом отмечается отсутствие половых различий в содержании мРНК андрогеновых рецепторов в латеральной перегородке, вентромедиальном ядре или аркуатном ядре [193]. Кроме того известно, что андрогены, регулируя ароматазную активность на уровне экспрессии гена в медиальной преоптической зоне и других компонентах мозга, опосредует половое поведение самцов. Так, для активации спаривания необходима ароматизация тестостерона в эстрадиол [233]. Это позволяет сделать предположение, что тестостерон и/или его эстрогенные метаболиты определяют в развивающемся мозге поло-специфичную способность к ароматизации. Они также вовлекаются в регулирование андрогенного ответа внутри компонентов нервной зоны, которые опосредуют половое поведение самцов [60, 166, 233].
Тестостерон способен оказывать влияние на синтез прогестерона in vitro [264].
Похожие работы

... нервной и эндокринной регуляции в организме // Усп. совр. биол. – 1981. – 19, № 3. – С. 380-382. 23. Вернигора А.Н., Бардинова Ж.С., Сметанин В.А., Генгин М.Т. Активность основных карбоксипептидаз в тканях самок крыс на разных стадиях эстрального цикла // Укр. биохим. журн. – 2003. – 75, № 5. – С. 99-102 24. Вернигора А.Н. Карбоксипептидаза Н мозга животных в норме и при действии стресс- ...
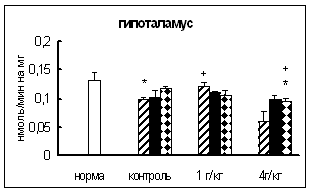
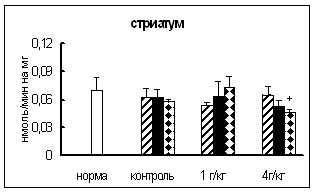
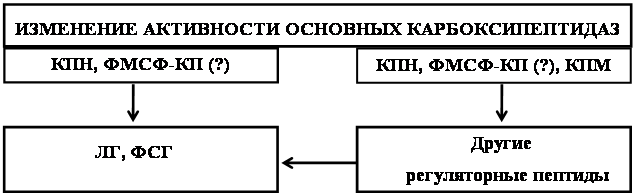
... о динамике изменения активности ферментов. ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ 3.1. Исследование активности основных карбоксипептидаз в тканях крыс разного возраста, испытавших пренатальное воздействие этанола 3.1.1. Исследование активности карбоксипептидазы Н в тканях пренатально алкоголизированных крыс разного возраста Согласно данным дисперсионного анализа пренатальное воздействие ...
0 комментариев