Навигация
Распространение дрожжевых грибов в природе
72371
знак
0
таблиц
0
изображений
2. Распространение дрожжевых грибов в природе
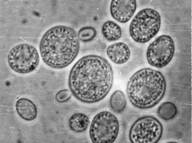
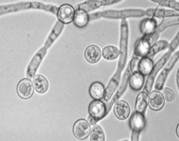
Особенности распространения дрожжей в природе стали интересовать микробиологов начиная с самых первых исследований процессов традиционного виноделия. Первоначально изучение дрожжей ограничивалось теми видами и штаммами, которые вызывали брожение при приготовлении пива и вина. Однако уже в конце XIX в. М. Бейеринк высказывал мысль о том, что эти культурные виды представляют собой селекционированные формы «диких» дрожжей, широко распространенных в природе. Естественно, возник вопрос об источниках их попадания в бродящие субстраты. Первые исследования, выполненные основателями зимологии Э. Хансеном и А. Клекером, были посвящены именно этой теме: поиску природных источников винных дрожжей Saccharomyces cerevisiae. В нашей стране этому вопросу также уделяли внимание крупнейшие микробиологи, например Г.А. Надсон. Сахаромицеты были найдены на ягодах винограда, однако, как оказалось, преобладают здесь совсем иные виды дрожжей, не участвующие в последующем сбраживании виноградного сока. Еще реже встречались сахаромицеты в окружающих субстратах, в частности, в почве под виноградниками. Уже в ранних работах высказывалось предположение, что почва не является средой, в которой возможно активное развитие дрожжевых грибов, а служит для последних лишь своеобразной «ловушкой», где дрожжи могут сохраняться определенное время в жизнеспособном состоянии и служить источником спор для инфицирования винограда нового урожая. Таким образом возникло понятие «круговорота дрожжей» в природе. Под «дрожжами» в то время подразумевались одноклеточные аскомицетовые грибы, родственные сахаромицетам и способные к активному брожению. . Расширяющиеся микологические исследования приводили к обнаружению все новых видов дрожжевых грибов, в том числе и таких, которые существенно отличались от типичных сахаромицетов. Оказалось, что многие из одноклеточных грибов, выделяемых из природных источников, не образовывали аскоспор и вообще не были способны к сбраживанию сахаров. Для таких дрожжей была создана серия формальных родов (Torulopsis, Candida, Rhodotorula, Cryptococcus), виды которых часто обнаруживаются в самых различных природных субстратах, включая почву, растения, разнообразные растительные остатки и природные воды. Стало понятным, что дрожжи распространены довольно широко, и их развитие далеко не ограничивается субстратами традиционных бродильных процессов. Однако, при этом существенно изменилось и содержание самого понятия «дрожжи». Дрожжами стали называть любые одноклеточные грибы, не обязательно вызывающие спиртовое брожение. В то же время, несмотря на существенные отличия между сахаромицетами и небродящими дрожжевыми грибами, достаточно долго сохранялось представление о дрожжах как самостоятельной филогенетической линии грибов. Лишь после обнаружения у несовершенных дрожжей рода Rhodotorula полного жизненного цикла, типичного для телиоспоровых гетеробазидиомицетов, термин «дрожжи» окончательно утратил таксономическое содержание. Тем не менее, вплоть до настоящего времени дрожжи продолжают рассматриваться в качестве единой группы, представляющей собой особую жизненную форму, или экоморфу грибов. Дело в том, что все дрожжи обладают очень сходным обликом за счет роста преимущественно в виде одиночных клеток. Кроме того, с одноклеточной организацией дрожжей сопряжены многие их физиологические особенности, в частности узкий спектр усваиваемых соединений, отсутствие способности к гидролизу труднодоступных полимеров, особенно таких, как целлюлоза и лигнин, быстрый рост за счет потребления простых углеводов. Особенно характерен этот набор признаков для аскомицетовых дрожжей. Все это делает их более приспособленными к обитанию в жидких и мелкодисперсных средах, богатых легкодоступными источниками углерода, в то время как мицелиальные грибы получают преимущество при росте на плотных поверхностях. Одноклеточность у грибов - вторичное явление в их эволюции, которое возникало независимо в разных группах аско- и базидиомицетов как реакция на существование в жидких и полужидких средах с относительно высокой концентрацией легкодоступных источников питания. Вскоре после открытия Пастером дрожжевой природы спиртового брожения было показано, что дрожжи постоянно обитают на поверхности ягод винограда и других сладких плодов, в цветочном нектаре, в кишечнике многих ксилофагов, проложенных ими галереях и личиночных камерах, в измельченной древесине, в кишечниках самых разных беспозвоночных, на наземных частях растений, в почве и т.д.
3. Жизненные формы дрожжейСпециализация на выполнении неодинаковых функций приводит у разных групп дрожжевых грибов к формированию характерного комплекса морфологических и физиологических свойств. Это дает возможность говорить о различных жизненных формах дрожжей. В общей экологии термином «жизненная форма» обозначается внешний облик, определенный морфологический тип организма, сформировавшиеся в результате приспособления к определенной среде обитания. Как уже отмечалось, дрожжи в современном понимании представляют собой определенную жизненную форму грибов. У микроорганизмов приспособления носят в основном физиологический характер, и при выделении таких экологических групп необходимо учитывать физиологические характеристики, поэтому правильнее говорить не о жизненных формах, а о морфо-физиологических группах. Среди дрожжей можно выделять следующие жизненные формы:
· Сахаробионты - «настоящие» дрожжи, наиболее типичным представителем которых является Saccharomyces cerevisiae. Они обладают комплексом свойств, свидетельствующим об их приспособленности к существованию в средах, обогащенных легкодоступными источниками углерода. Отсутствие пигментации, развитых мицелиальных структур, хламидоспор, слизистых капсул, а также способность к более или менее интенсивному брожению и узкий спектр усваиваемых соединений углерода - характерный набор свойств этих дрожжей. Кроме сахаромицетов к сахаробионтам следует относить представителей родов Debaryomyces, Kluyveromyces, Torulaspora, Zygosaccharomyces, а также большинство видов из родов Pichia и Candida.
· Фитобионты - адаптированы к обитанию на поверхности живых частей растений и, как правило, образуют каротиноидные пигменты. Они часто имеют в цикле развития хламидоспоры или хламидоспороподобные клетки, устойчивые к высушиванию. Характерный признак многих видов - образование баллистоспор, рассеивающихся токами воздуха. Наиболее типичные представители фитобионтов - роды Sporobolomyces и Sporidiobolus, некоторые виды родов Rhodotorula и Cryptococcus.
· Сапробионты обладают относительно высокой гидролитической активностью и принимают участие в деструкции растительных остатков на средних и поздних стадиях. К типичным сапробионтам относятся некоторые виды рода Trichosporon, Cystofilobasidium capitatum, группа несовершенных видов базидиомицетового аффинитета, классифицируемых в роде Cryptococcus (Cryptococcus podzolicus, Cryptococcus humicolus).
· Педобионты - дрожжи, наиболее приспособленные к обитанию на твердых поверхностях почвенных частиц. Они обладают слизистыми капсулами, которые создают межклеточную среду, сохраняющую благоприятный режим влагообмена и питания в условиях временного иссушения почвы. Эти дрожжи способны накапливать большое количество запасных веществ, главным образом в форме липидов, которые обеспечивают переживание длительных периодов голодания. Для них также характерна способность к усвоению соединений азота в очень низкой концентрации. Типичные представители педобионтов - все виды липомицетов. По-видимому, к педобионтам можно также отнести некоторые виды криптококков, в частности Cryptococcus terreus, Cryptococcus aerius, Cryptococcus terricola.
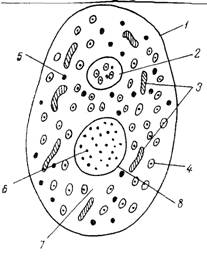
4. Дрожжевая клетка. ЦитологияДожжевая клетка имеет все основные структуры, которые присущи любой эукариотической клетке, но в то же время она обладает особенностями, свойственными грибам, а именно, сочетанием признаков как растительной, так и животной клеток: клеточная стенка у них ригидная, как у растений, но в клетке отсутствуют хлоропласты и накапливается гликоген, как у животных.
Компоненты дрожжевой клетки
Ядро
В дрожжевой клетке в фазе между делениями всегда имеется только одно ядро. В световом микроскопе его можно увидеть после специальной окраски или с помощью фазово-контрастного устройства при высоких разрешениях. На электронно-микроскопических снимках ультратонких срезов дрожжевых клеток ядро выглядит как более или менее округлая органелла, окруженная двойной мембраной. В ней есть поры в виде округлых сквозных отверстий, которые образуются в результате слияния двух ядерных мембран. Однако ядерные поры - не просто отверстия, они заполнены сложноорганизованными структурами, которые называют комплексом пор ядра. Считается, что основная функция ядерных пор - транспорт готовых рибосомных субъединиц в цитоплазму. Ядерная оболочка многофункциональна, но в основном играет роль барьера, отделяющего содержимое ядра и регулирующего транспорт макромолекул между ядром и цитоплазмой. Основные функциональные единицы ядра - молекулы ДНК, несущие основную генетическую информацию о клетке. ДНК составляет основную часть хроматина - основного компонента ядра. Число хромосом в ядре разных видов дрожжей может быть различным, оно колеблется от 2 до 16.
Митохондрии
В митохондриях имеется собственная митохондриальная ДНК (мДНК), а также весь аппарат белкового синтеза, включая матричную РНК и 70S рибосомы (в отличие от 80S рибосом в цитоплазме). мДНК у дрожжей составляет 5-20% от всей ДНК клетки. Число митохондрий в одной дрожжевой клетке варьирует в пределах 1-20 в разные периоды роста и в зависимости от условий. Как правило, 1-2 митохондрии в клетке более крупные, чем остальные и имеют разветвленную форму. Реконструкция ультратонких срезов клетки позволяет предположить, что в некоторых случаях (в подготовительный период почкования) клетка содержит всего одну вытянутую и сильно разветвленную митохондрию. Митохондрии способны к самовоспроизведению.
Цитоплазаматическая мембрана
На поперечном срезе под электронным микроскопом мембрана у дрожжей выглядит как трехслойная структура. Она представляет собой два слоя фосфолипидов, в которые погружены белковые молекулы, то есть построена по общему для всех клеточных мембран принципу. Однако, имеются различия, касающиеся химического состава. У Saccharomyces cerevisiae основными фосфолипидами мембран являются лецитин, фосфатидилэтаноламин и фосфатидилсерин. На их долю приходится около 90% всех липидов мембраны. В состав мембраны дрожжей входят стероиды - эргостерол, зимостерол и др. Белки представлены в основном ферментами, которые участвуют в трансмембранном переносе веществ, расщеплении полисахаридов и синтезе внеклеточных структур. . Функции цитоплазматической мембраны многообразны: регуляция биосинтеза клеточной стенки, активный транспорт транспорт в клетку специфических молекул органических веществ, транспорт ионов K+ и Na+ и др.
Вакуоли
В фазово-контрастном микроскопе в клетках дрожжей хорошо видны светлые и прозрачные структуры круглой формы. Это вакуоли (см. приложение 3). Обычно их 1-3 в клетке. Каждая вакуоль окружена одинарной мембраной и содержит различные ферменты, липиды, низкомолекулярные продукты метаболизма (аминокислоты), ионы металлов. В вакуолях сосредоточена большая часть ионов калия. Иногда в вакуоли видны «пляшущие» за счет броуновского движения плотные гранулы. Это так называемые метахроматические гранулы, «пляшущие тельца» (dancing bodies), или волютин. Гранулы эти состоят из полимеризованных остатков фосфатов, а по периферии они покрыты комплексными соединениями из РНК, белков и липидов. Волютин - это резерв полифосфатов в клетке. Основная функция вакуолей - разобщение процессов синтеза и распада белков и нуклеиновых кислот. Они выполняют также роль депо для хранения некоторых запасных веществ и ферментов, участвуют в регуляции тургорного давления. Также в клетке присутствуют: клеточная стенка, которая защищает протопласт от осмотического разрыва и придает клетке определенную форму; капсула (слизистый полисахаридный чехол вокруг клетки), цитоплазма и липиды.
Похожие работы
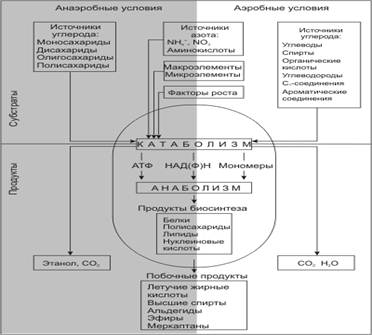
... так как от их состава зависит качество пищевого продукта, получаемого с помощью дрожжей. Такие соединения называют органолептическими. Обязательные побочные продукты метаболизма дрожжей – высшие спирты (сивушные масла). Механизм синтеза высших спиртов связан с образованием алифатических аминокислот. Чаще всего встречаются спирты: пропиловый, изоамиловый, бутиловый, изобутиловый. Летучие жирные ...

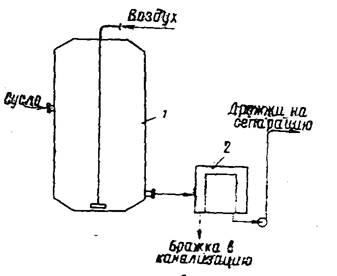
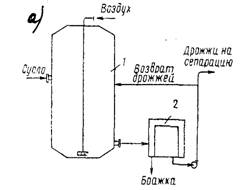
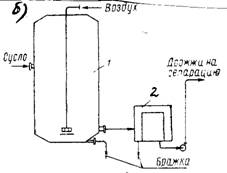
... часть фосфорной кислоты связана в дрожжах с органическими соединениями. Общее количество Р2О5 у сахаромицетов колеблется в пределах от 3,2 до 4,4% к сухому веществу. 2 Технология производства сухих дрожжей 2.1 Этапы производства дрожжей В процессе выращивания дрожжей из одной клетки получают несколько тонн продукта. Начальная стадия выращивания проходит в микробиологической лаборатории. ...
... в качестве объектов исследования типичные и луговые степи Русской равнины, а также зоомикробные комплексы диплопод, питающихся растительным опадом. Были изучены численность и таксономическая структура блока сапротрофных бактерий и дрожжей в разных ярусах степных биогеоценозов (БГЦ) и в пищевых цепях диплопод (Черняковская и др., 1990; Вино-варова, 1989; Byzov, et al.,1996). Было установлено, что ...
... любого, кто украдет росток, чтобы его возделывали в другой стране.[11] Учитывая все вышесказанное, утверждать с полной уверенностью - так сказать, однозначно, кто именно и когда первым стал использовать в технологии пивоварения хмель, весьма затруднительно. Но это, впрочем, не так уж важно. Главное то, что в результате у пива появился тот "пивной" вкус, который мы хорошо знаем сегодня.[11] Что ...
0 комментариев