Навигация
Половое размножение и жизненные циклы дрожжей
72371
знак
0
таблиц
0
изображений
6. Половое размножение и жизненные циклы дрожжей
Половое размножение - это сложная цепь событий, включающая контакт двух гаплоидных клеток, их слияние (сначала слияние цитоплазмы - плазмогамия, а затем сразу же или со значительной задержкой слияние ядер - кариогамия), образование диплоидной зиготы, ядро которой затем либо делится мейотически с восстановлением гаплоидного состояния, либо дает начало диплоидному поколению клеток. Таким образом, половое размножение связано со сменой ядерных фаз. У дрожжей, как и у всех грибов, чередование ядерных фаз сопряжено с образованием половых гаплоидных спор – аскоспор (см. приложение 12) или базидиоспор (см. приложение 13). Весь ход событий в развитии организма от одной стадии до этой же стадии в следующем поколении составляет жизненный цикл, или онтогенез. Полный жизненный цикл включает вегетативную стадию, в течение которой клетки размножаются при помощи митотического деления, и половой цикл, включающий мейотическое деление ядра. У мицелиальных грибов вегетативное размножение занимает обычно одну из стадий жизненного цикла: у аскомицетов - преимущественно гаплоидную, у базидиомицетов - дикариотическую. У дрожжей вегетативное размножение может происходить в любой фазе жизненного цикла. На этом основании различают гаплоидные виды, у которых вегетативное размножение происходит в гаплоидной фазе, диплоидные виды, размножающиеся вегетативно в диплофазе, а также гапло-диплоидные, образующие стабильные как гапло-, так и диплофазы, или же смешанные популяции гаплоидных и диплоидных клеток. У дрожжей встречаются различные типы полового процесса. У большинства видов в половом процессе у дрожжей участвуют обычные соматические, то есть неспециализированные клетки. Такой тип полового процесса называется соматогамией. Существуют разновидности соматогамии:
· гологамия - слияние (копуляция) двух морфологически сходных соматических клеток,
· педогамия - слияние материнской и дочерней клетки-почки,
· адельфогамия - слияние сестринских клеток-почек.
· У некоторых видов аскомицетовых дрожжеподобных грибов половой процесс представляет собой типичную для большинства аскомицетов гаметангиогамию - копуляцию специальных клеток мицелия - гаметангиев. Копулировать могут не любые две клетки, а лишь клетки, относящиеся к различным типам спаривания.
Термин «тип спаривания» используется вместо термина «пол» в том случае, когда копулирующие клетки не имеют морфологических отличий, а различаются лишь физиологически. У одних дрожжей при вегетативном размножении происходит разделение клеток на различные типы спаривания (обозначаемые a и α), и, следовательно, в потомстве одной клетки возможен половой процесс. Такие дрожжи называют гомоталличными. У других клетки не способны переключаться с одного типа спаривания на другой, и половой процесс в потомстве одной клетки невозможен. В этом случае дрожжи называют гетероталличными. Половой процесс у таких дрожжей происходит только при объединении клеток из популяций a и α типов спаривания.
Дифференциация пола у дрожжей
Особенности половой дифференциации и конъюгации клеток при половом размножении наиболее хорошо изучены у дрожжей Saccharomyces cerevisiae. При половом размножении могут конъюгировать не любые две клетки, а только клетки различных типов спаривания. Эти клетки различаются между собой только по одному генетическому локусу, обозначаемому mat (mating - спаривание). Локус mat может находиться в двух аллельных состояниях: mat a и mat α. Клетки, несущие локус mat a или mat α, обозначаются соответственно как a- и α-клетки. Локусы mat a и mat α ответственны за образование так называемых половых факторов (соответственно a- и α-фактора), которые представляют собой олигопептиды. a-Фактор, образуемый а-клетками, способен блокировать процесс митотического деления у α-клеток. α-Фактор, образуемый α-клетками, блокирует митотическое размножение у а-клеток. Поэтому конъюгация возможна только при взаимодействии а- и α-клеток, но не а х a или α х α. После конъюгации а- и α-клеток образуется диплоидная клетка (зигота), имеющая генотип а/α. Только такие клетки способны к мейотическому делению с образованием половых спор. В процессе мейотического деления а/α клетки образуется 4 гаплоидных ядра, два из которых а-типа, а два - α-типа. Поэтому в типичном четырехспоровом аске две аскоспоры принадлежат к а-типу, а две - к α-типу. Более сложные случаи генетической детерминации пола наблюдаются у базидиальных дрожжей. У видов в родах Rhodosporidium, Leucosporidium, Sporidiobolus тип спаривания не биполярный, а тетраполярный, так как определяется не двумя (а и α), а четырьмя аллелями, которые принято обозначать как А, а, В, в. В этом случае скрещивание возможно лишь между штаммами, которые различаются всеми четырьмя аллелями: АВ ґ ав или Ав ґ аВ. В комбинациях ав ґ ав или АВ ґ АВ спаривание невозможно. При взаимодействии штаммов, различающихся только по одному локусу: АВ ґ аВ, АВ ґ Ав, ав ґ Ав или ав ґ аВ, иногда наблюдается копуляция клеток, но дикариотический мицелий, как правило, не развивается.
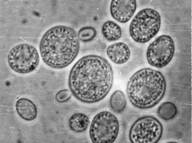
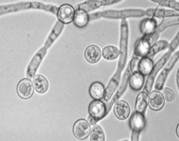
Аскомицетовые дрожжиК аскомицетам относят дрожжи, половые споры которых формируются эндогенно внутри особых вместилищ - сумок, или асков. В отличие от мицелиальных аскомицетов, большая часть жизненного цикла которых проходит в гаплоидной фазе, а диплоидна только молодая сумка, среди аскоспоровых дрожжей есть виды с разными типами онтогенеза: гаплоидным, диплоидным и гапло-диплоидным. При этом аском может стать либо непосредственно зигота (Schizosaccharomyce, Zygosaccharomyces), либо отдельная диплоидная вегетативная клетка (Saccharomyces, Saccharomycodes), либо сумка развивается как новообразование с участием так называемых «активных» почек, выполняющих функцию гамет (Lipomyces). Аски могут также формироваться из хламидоспор (см. приложение 11), из клеток псевдомицелия (см. приложение 8), или на истинном мицелии. В любом случае формирование аска происходит из диплоидной клетки после мейотического деления ядра. Мейоз представляет собой два следующих друг за другом деления ядра с однократным удвоением хромосом. Перед вторым делением хромосомы не удваиваются, поэтому в результате происходит редукция числа хромосом и возникают четыре гаплоидных ядра. Затем в районе ядерных бляшек начинают формироваться мембраны, которые постепенно разрастаются и охватывают часть цитоплазмы будущего аска. После образования аскоспор остается небольшое количество «неиспользованной» цитоплазмы. Между двумя листками мембран закладывается клеточная стенка аскоспоры. Аскоспоры отличаются от вегетативных клеток более толстой и многослойной клеточной стенкой, меньшим развитием внутриклеточных мембранных систем, отсутствием вакуолей. Эти особенности связаны с пониженной метаболической активностью аскоспор. Обычно аскоспоры содержат большое количество запасных веществ, чаще всего - липидов. Аскоспоры дрожжей - это типичные покоящиеся споры, способные более или менее длительный период существовать в неактивном состоянии. Однако, устойчивость аскоспор к ряду повреждающих факторов, например, к повышенным температурам, обычно не намного выше, чем у вегетативных клеток. Форма аскоспор у дрожжей очень разнообразна. Они могут быть круглыми, овальными, бобовидными, чечевицеобразными, серповидными, игловидными и т.д. Кроме того, аскоспоры могут иметь на поверхности клеточной стенки различные скульптурные образования, которые хорошо выявляются в электронном микроскопе. За счет таких образований аскоспоры могут быть бородавчатыми, сатурновидными, напоминающими грецкий орех и пр. Морфология асков зависит от способа их образования, типа полового процесса (гологамия или педогамия), количества и формы аскоспор. Количество аскоспор в аске у разных видов может быть от одной до нескольких десятков. Наиболее часто встречаются аски с 1,2,4 и 8 аскоспорами. Аски различаются также временем существования. У большинства дрожжей аски устойчивые и разрушаются только в очень старых культурах. Но есть виды с быстро разрушающимися асками. Морфология асков, аскоспор, тип полового процесса имеют большое таксономическое значение. Эти признаки используются в систематике дрожжей при выделении таксонов родового уровня. . Рассмотрим несколько примеров жизненных циклов аскомицетовых дрожжей.
· Гаплоидные. У таких дрожжей вегетативное размножение происходит в гаплоидной фазе, а диплоидная стадия очень короткая: образовавшееся диплоидное ядро сразу же делится мейотически с восстановлением гаплоидного состояния. Schizosaccharomyces pombe. Половой процесс - гологамия. Две морфологически сходные гаплоидные вегетативные клетки образуют выросты, с помощью которых происходит контакт, а затем слияние содержимого клеток. Возникает диплоидная зигота, которая вегетативно не размножается, а ядро ее сразу переходит к мейотическому делению. Образующиеся четыре гаплоидных ядра включаются в аскоспоры. После освобождения из аска аскоспоры прорастают и дают начало длительной стабильной вегетативной фазе. Lipomyces tetrasporus. Половой процесс - адельфогамия. Роль гамет здесь выполняют активные почки. Такие почки формируются на гаплоидных вегетативных клетках на поздних стадиях роста после периода вегетативного размножения. Обычно две почки на одной материнской клетке выполняют функцию гамет и копулируют между собой, образуя зиготу, которая затем разрастается в виде мешка и отделяется перегородкой от несущей ее клетки. Ядро зиготы делится мейотически и она превращается в четырехспоровый мешковидный аск, прикрепленный к материнской клетке, на которой может затем формироваться вторая и третья сумки.
· Диплоидные. У этих дрожжей вегетативно размножаются только диплоидные клетки. Гаплофаза ограничена молодыми асками и аскоспорами. . Saccharomycodes ludwigii. У этих дрожжей диплоидизация происходит при слиянии аскоспор. Четыре гаплоидные аскоспоры прорастают и начинают копулировать попарно, когда они еще находятся в аске. Образовавшиеся диплоидные клетки размножаются вегетативно, образуя стабильную и длительную диплофазу. При соответствующих условиях, когда снимается контроль митотического деления ядра, диплоидная клетка вступает в митотический цикл и превращается в аск с 4 аскоспорами. Hanseniaspora uvarum. Жизненный цикл сходен с описанным выше за исключением того, что споры не копулируют, но ядро в зрелой споре после освобождения ее из сумки в условиях, обеспечивающих вегетацию, делится мейотически. Образовавшиеся два гаплоидных ядра сливаются, образуя уже диплоидную клетку, способную к вегетативному размножению. Сходный цикл наблюдается и у почвенных дрожжей Williopsis saturnus.
· Гапло-диплоидные. Существуют дрожжи, у которых вегетативное размножение может происходить как в гаплоидной, так и в диплоидной фазах. Длительность той или другой фазы зависит от вида и от условий роста. Saccharomyces cerevisiae. Эти дрожжи вегетируют преимущественно в диплоидном состоянии, но у них имеется короткая вегетативная гаплоидная фаза. Диплоидная клетка в условиях дефицита легкодоступных источников углерода прекращает почковаться, и ядро ее делится мейотически. В результате она превращается в аск с 4 гаплоидными аскоспорами, которые после освобождения из аска прорастают и образуют гаплоидное поколение. Гаплоидные клетки обычно мельче диплоидных и имеют более округлую форму. Шрамы почкования у них сближены, почки образуются группами в одном локусе. После нескольких циклов почкования две клетки конъюгируют и сливаются, восстанавливая диплоидное состояние.
Базидиомицетовые дрожжи
В отличие от аскоспоровых дрожжей, все дрожжи, которые относят к базидиомицетам, образуют в ходе полового размножения специальные клетки - базидии, несущие экзогенные споры (базидиоспоры). Значительную часть жизненного цикла занимает дикариотическая фаза, представленная дикариотическим мицелием с пряжками. Большинство видов гетероталличны и поэтому многие из них долгое время были известны только в гаплофазе как несовершенные дрожжи. Жизненные циклы базидиомицетовых дрожжей также достаточно разнообразны. Активное исследование жизненных циклов базидиомицетовых дрожжей началось лишь в конце 60-х - начале 70-х годов XX в., после того как японский миколог Исао Банно опубликовал результаты открытия полового цикла у хорошо известных и ранее считавшихся несовершенными красных дрожжей Rhodotorula glutinis. Ему удалось подобрать типы спаривания среди большого числа проверенных штаммов, и в результате их скрещивания он наблюдал образование дикариотического мицелия, на котором формировались структуры, морфологически сходные с хламидоспорами, то есть крупные клетки с сильно утолщенными оболочками и большим запасом внутриклеточных липидов. Именно в них затем происходило слияние двух ядер и последующий мейоз с восстановлением гаплоидного состояния. Эти клетки получили название телиоспор. При прорастании они образовывали промицелий с поперечными септами, делящими его на 4 клетки. Эти клетки отпочковывали гаплоидные споридии. Совершенная стадия Rhodotorula glutinis была названа Банно Rhodosporidium toruloides. Позже сходный цикл был описан для некоторых баллистоспоровых дрожжей и для нескольких видов несовершенного рода Candida. Их телиоспоровые стадии получили соответственно родовые названия Sporidiobolus и Leucosporidium. Сейчас похожие полные или не завершенные жизненные циклы известны для многих базидиомицетовых дрожжей. Они различаются формой телиоспор, характером их прорастания и морфологией базидий. . Другой тип жизненного цикла базидиомицетов с дрожжевой анаморфой известен для дрожалковых грибов порядка Tremellales. Эти грибы известны достаточно давно, так как в онтогенезе этих организмов есть стадия образования макроскопических плодовых тел, которые в природе обычно развиваются на старой древесине, на стволах мертвых деревьев. То, что базидиоспоры дрожалковых грибов способны к почкованию, было замечено микологами еще в конце XIX в., однако наличие самостоятельной дрожжевой фазы в цикле развития тремелловых было установлено после детального изучения видов рода Tremella в 1960-х гг. В культурах отдельные базидиоспоры гетероталличных видов дают начало дрожжевым стадиям. Мицелий обычно развивается только при смешивании штаммов совместимых типов спаривания. Смешение соответствующих типов спаривания приводит к быстрой реакции клеток на выделяющиеся феромоны; почкование прекращается, развиваются конъюгационные трубки, через которые клетки копулируют, происходит плазмогамия и развивается мицелиальная дикариотическая стадия. У многих видов в подходящих условиях вскоре после формирования дикариотического мицелия начинается развитие плодовых тел. В плодовых телах образуются 2-4-клеточные гетеробазидии. Освободившиеся базидиоспоры могут размножаться вегетативно почкованием или формированием баллистоспор. Характеристики жизненного цикла играют большую роль в систематике базидиомицетовых дрожжей. Однако очень часто у базидиомицетовых дрожжей, изолируемых из природных местообитаний, не удается наблюдать полный жизненный цикл при культивировании на лабораторных средах, поэтому их идентификация и классификация вызывают большие трудности. Значительные успехи в систематике таких анаморфных базидиомицетовых дрожжей достигнуты в последние десятилетия, особенно в связи с внедрением в таксономическую практику молекулярно-биологических методов. Как оказалось, группирование базидиомицетовых дрожжей на основе молекулярно-биологических признаков, в частности нуклеотидных последовательностей рРНК, часто не совпадает с характеристиками их жизненного цикла, особенно такими, как способность к образованию телиоспор, морфология базидий. Поэтому классификация базидиомицетовых дрожжей еще далека от завершенности и требует дальнейших исследований.
Похожие работы
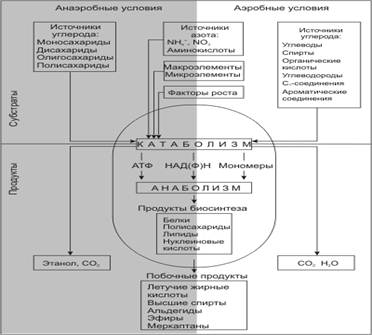
... так как от их состава зависит качество пищевого продукта, получаемого с помощью дрожжей. Такие соединения называют органолептическими. Обязательные побочные продукты метаболизма дрожжей – высшие спирты (сивушные масла). Механизм синтеза высших спиртов связан с образованием алифатических аминокислот. Чаще всего встречаются спирты: пропиловый, изоамиловый, бутиловый, изобутиловый. Летучие жирные ...

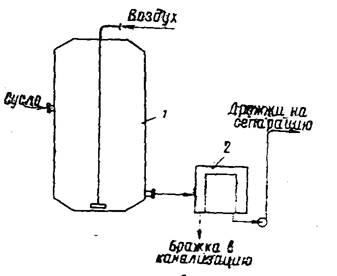
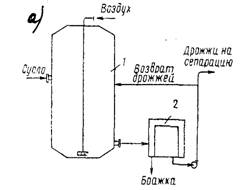
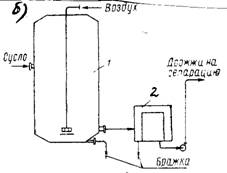
... часть фосфорной кислоты связана в дрожжах с органическими соединениями. Общее количество Р2О5 у сахаромицетов колеблется в пределах от 3,2 до 4,4% к сухому веществу. 2 Технология производства сухих дрожжей 2.1 Этапы производства дрожжей В процессе выращивания дрожжей из одной клетки получают несколько тонн продукта. Начальная стадия выращивания проходит в микробиологической лаборатории. ...
... в качестве объектов исследования типичные и луговые степи Русской равнины, а также зоомикробные комплексы диплопод, питающихся растительным опадом. Были изучены численность и таксономическая структура блока сапротрофных бактерий и дрожжей в разных ярусах степных биогеоценозов (БГЦ) и в пищевых цепях диплопод (Черняковская и др., 1990; Вино-варова, 1989; Byzov, et al.,1996). Было установлено, что ...
... любого, кто украдет росток, чтобы его возделывали в другой стране.[11] Учитывая все вышесказанное, утверждать с полной уверенностью - так сказать, однозначно, кто именно и когда первым стал использовать в технологии пивоварения хмель, весьма затруднительно. Но это, впрочем, не так уж важно. Главное то, что в результате у пива появился тот "пивной" вкус, который мы хорошо знаем сегодня.[11] Что ...
0 комментариев