Навигация
Технология получения глутамата
26888
знаков
2
таблицы
9
изображений
3.5. Технология получения глутамата.
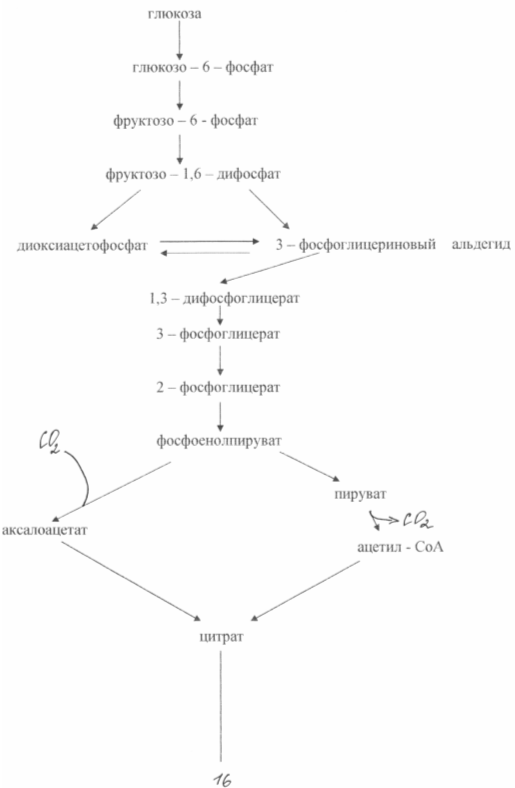
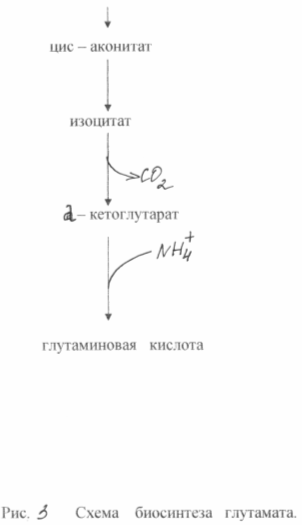
В основе сверхсинтеза глутаминовой кислоты из глюкозы у этих бактерий лежат два биохимических принципа: недостаток фермента а - кетоглутаратдегидрогеназы и блокировка биосинтеза биотина. Неспособность клеток синтезировать биотин приводит к увеличению проницаемости цитоплазмотической мембраны, что повышает экскрецию глутамата. Он образуется в результате аминирования а - кетоглутарата, неспособного к дальнейшим превращениям в цикле трикарбоновых кислот. Схема биосинтеза глутамата из глюкозы у данного типа мутантов показана на рис. 3
При биосинтезе глутаминовой кислоты очень большое значение имеет концентрация биотина в среде. Необходимо обеспечить его концентрацию 1 - 5 мкг /л. В этом случае нарушается нормальный синтез фосфолипидов мембраны и последняя становится проницаемой для глутамата. При концентрации биотина 15 мкг/л наблюдается интенсивный рост биомассы. Проницаемость цитоплазмотической мембраны для глутамата можно снизить также при помощи пенициллина, добавляя его к среде во время логарифмической фазы роста. В этом случае фосфолипиды экстрагируются из мембраны и транспорт глутамата может осуществляться в течение 40 - 50 часов. Бактериальный синтез глутамата позволяет получать примерно 50 % - ный выход продукта из сахара и накапливать в среде ферментации до 200 г/л глутамата. Известны методы получения глутамата на этанольных средах ( до 60 г/л ) или на ацетате (до 98 г/ л ).[6]
4. ПРОМЫШЛЕННЫЙ СИНТЕЗ АМИНОКИСЛОТ
Промышленное производство аминокислот осуществляется двумя способами: микробиологическим и химическим.
4.1 Микробиологический синтез
Микробиологический синтез основан на выращивании определенных видов микроорганизмов на питательных средах, имеющих подходящий источник углерода. Чаще всего это сахара, содержащиеся, например, в патоке. Мутированные микроорганизмы с нарушенным азотным обменом выделяют в раствор большое количество какой-либо одной аминокислоты. После окончания процесса ферментации аминокислоту выделяют из раствора химическими методами
Путем микробиологической ферментации получают основное количество глутаминовой кислоты и весь лизин. У этого процесса свои преимущества и свои недостатки. С одной стороны, в нем мало стадий и требуется относительно простая и универсальная аппаратура. С другой стороны, живые микроорганизмы, с которыми приходится работать, очень чувствительны к малейшему изменению условий, а концентрация целевого продукта получается низкой, что ведет к увеличению размеров аппаратуры.
Существует способ микробиологического получения фенилаланина при помощи тирозин - и метиониндефицитного мутанта Brevibacterium lactofermentum. В периодическом процессе ферментации достигнута концентрация продукта 24,8 г/л. Однако для данного процесса требуются сложные и дорогие среды. Определенный интерес представляют биосинтез фенилаланина ауксотрофным мутантом Е. coli, который можно культивировать в глюкозной среде с фосфатами. Процесс ферментации осуществляют доливным методом с рециркуляцией биомассы. Биомасса в реакторе 60 - му часу достигает 45 - 50 г/л, а концентрация фенилаланина - 22,4 - 22,8 г/л. Продуктивность системы 0,72-0,86 г/( лч ); выход продукта 0,11г.
4.2 Химический синтез
Химический синтез более универсален, чем микробиологический, и позволяет получать соединения любой возможной структуры. Здесь используется непищевое минеральное сырье, достигается любая концентрация продукта, однако, как правило, процесс многостадиен и требует более сложной аппаратуры.
Оба способа обеспечивают получение природных аминокислот необходимой степени химической и оптической чистоты. Так что в конечном счете, когда речь идет о промышленном производстве, последнее слово остается за экономикой: по данным зарубежных специалистов, при больших масштабах химические методы становятся более рентабельными.
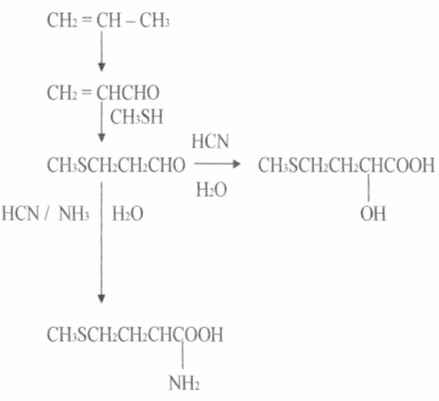
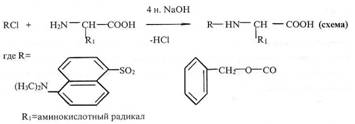
Наиболее широко разработан промышленный синтез метионина- аминокислоты, главным потребителем которой является птицеводство. Исходным веществом служит пропилен - продукт крекинга нефти. Пропилен окисляется до акролеина, который в результате серии реакций, превращается в рацемический метионин.
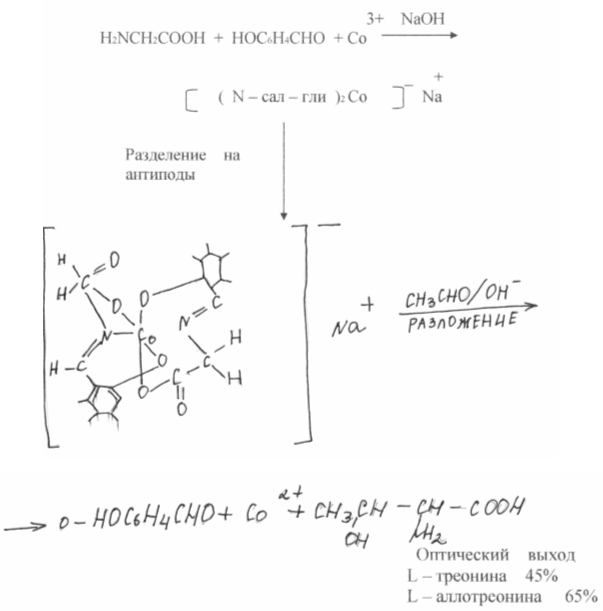
В результате химического синтеза обычно получается смесь равных количеств L и D - изомеров аминокислот, в то время как в состав белков входят исключительно L-изомеры. Эти же изомеры питательны. D-изомеры организмом, как правило, не усваиваются и являются балластом. Следовательно, необходимо разделение, что неминуемо отрицательно сказывается на экономике. В последнее время в области расщепления рацемических смесей аминокислот достигнуты серьезные успехи. В работах СВ. Рогожина и В.А. Даванкова показано, что оптически неактивные аминокислоты, будучи ковалентно присоединены к нерастворимому полимерному носителю, легко образуют комплексы с медью, никелем и т.п. другая рацемическая аминокислота, находящаяся в растворе, занимает два вакантных координационных места у атома металла, причем прочность комплексов L - и D - изомеров различна. Сколь ни мало это различие, будучи повторенным многократно в процессе хромотографии, оно обеспечивает полное или частичное разделение оптических антиподов. Наилучшие результаты получены с DL - пролином, который может быть препаративно разделен на оптические изомеры.
Усилие многих исследователей направлены также на разработку такого химического синтеза, который давал бы только один желаемый природный оптически активный изомер - изомер, синтезируемый живой природой, - ассиметрического синтеза. В этом направлении за последние годы достигнуты серьезные успехи. В работах А. Кагана (Франция ) и Е, Корна ( США ) достигнуты практически количественные оптические выходы. Чрезвычайно заманчивым представляется воспроизведение путей синтеза аминокислот природными ферментными системами. Большое количество таких синтезов осуществляется пироксальзависимыми ферментами, причем сразу получается нужный оптический изомер аминокислоты. Большой вклад в изучение этих процессов сделан академиком А.Е. Браунштейном (Россия) и профессором Ю. Снеллом (США).
Российские ученые поставили своей целью найти химические системы, которые могли бы не только моделировать биохимические реакции, но и осуществлять процессы, не проходящие в живом организме. В качестве такой системы были выбраны комплексы шиффовых оснований аминокислот с ионами переходных металлов. Предпологалось, что салициловый альдегид в этих комплексах будет играть роль лиридоксаля, увеличивая реакционную способность С - Н - связи аминокислоты, ион металла будет делать тоже самое, но еще удерживать систему в жестком плоском состоянии, что в природе обеспечивает белок фермента. Общая диссимметрия комплекса позволяла надеяться, что в природе обеспечивает белок фермента. Общая диссиметрия комплекса позволяла надеяться, что реакция может быть проведена стереоспепифически, т.е. в результате дать оптически активную кислоту.
Такие реакции в природе не идут, а практическая ценность их заключается в том, что при этом сразу получается глутаминовая кислота. Таким образом, открывается путь нового общего синтеза аминокислот, проходящего в чрезвычайных условиях.
Несравненно сложнее оказатось воспроизвести другую сторону действия природных ферментов - ассиметричеекий синтез. Для этого комплексы были разделены на оптически активные антиподы.
Похожие работы
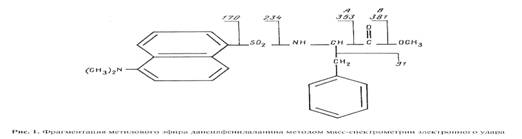
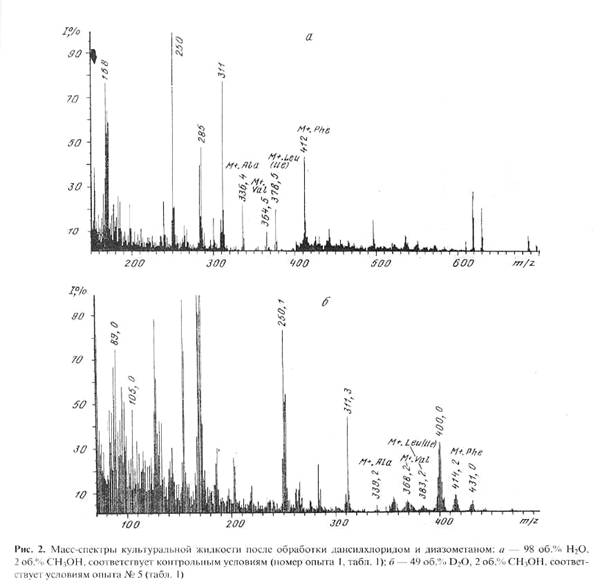
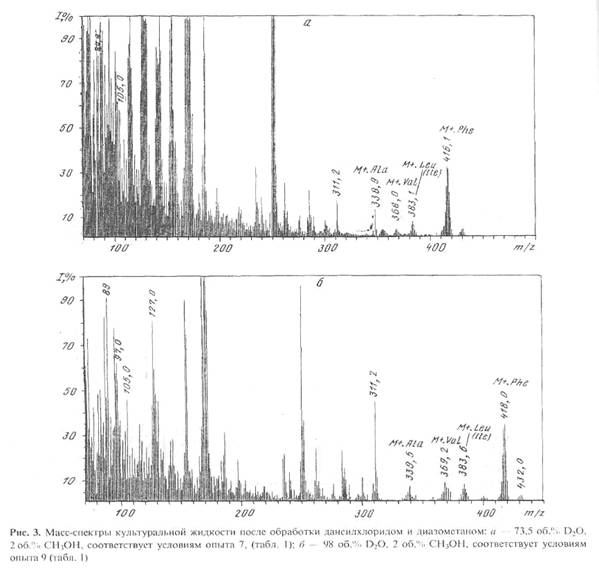
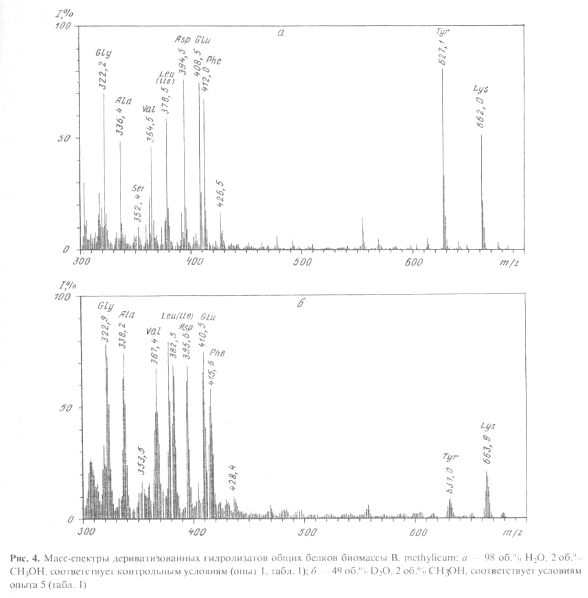
... использование данного штамма метилотрофных бактерий для наработки униформно меченного белка и аминокислот, было необходимо изучить уровни включения дейтерия в аминокислоты белковых гидролизатов B. methylicum, при росте бактерий на среде, содержащей максимальные концентрации тяжелой воды. Данные по степеням включения дейтерия в аминокислоты белка B. methylicum, полученного со среды, содержащей 98 ...
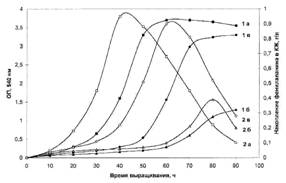
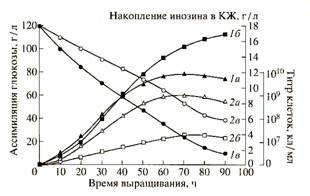
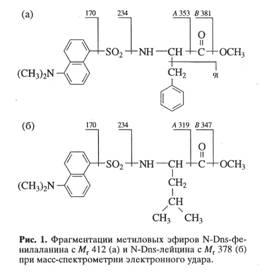
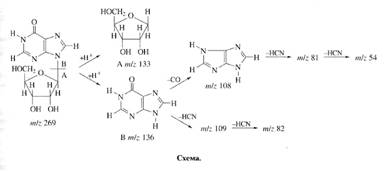
... . Так, в Phe детектировалось 80,5 % метки, в Ala - 95 %, в Gly - 90% (см. табл. 6). Во всех экспериментах степени включения дейтерия и изотопа углерода 13С в метаболически связанных аминокислотах обнаружили определённую коррелляцию. Так, степени изотопного обогащения валина и лейцина (семейство пирувата), фснилаланина и тирозина (семейство ароматических аминокислот} совпадают (табл. 6). Степени ...
... трансаминирования. Образуется альдегид и новая аминокислота, соответствующая исходной кетокислоте. Декарбоксилирование, связанное с реакцией конденсацией двух молекул: Обезвреживание аммиака в организме. В организме человека подвергается распаду около 70г аминокислот в сутки: при этом освобождается большое количество аммиака, являющегося высокотоксичным соединением. Поэтому крнцентрация аммиака ...
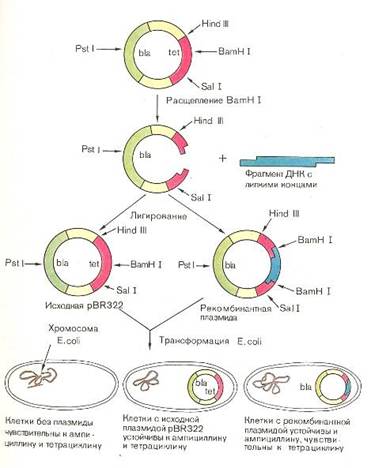
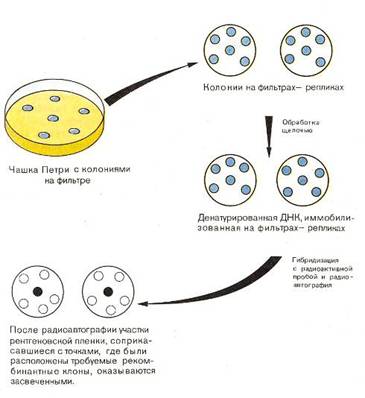
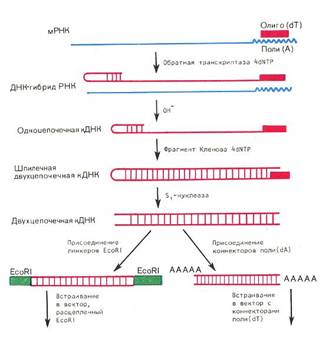
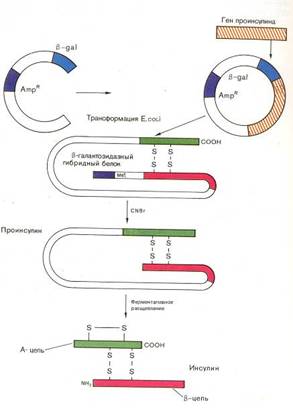
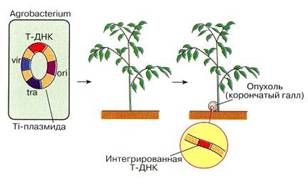
... инженерию. Необходимо отметить, что если базовый стандарт по химии не предусматривает изучение вопросов биотехнологии, то таковой по биологии содержит наиболее общие её аспекты: достижения генной инженерии и перспективы биотехнологии. 2.2 Межпредметные связи по изучению аспектов биотехнологии в средней школе По программе Р.Г. Ивановой и Л.А. Цветкова в 10 классе предусмотрено изучение темы ...
0 комментариев