Навигация
ПОЛУЧЕНИЕ ШТАММОВ-ПРОДУЦЕНТОВ БАС, АДАПТИРОВАННЫХ К
55634
знака
6
таблиц
5
изображений
1. ПОЛУЧЕНИЕ ШТАММОВ-ПРОДУЦЕНТОВ БАС, АДАПТИРОВАННЫХ К
РОСТУ И БИОСИНТЕЗУ НА СРЕДАХ С МАКСИМАЛЬНЫМИ
КОНЦЕНТРАЦИЯМИ ТЯЖЁЛОЙ ВОДЫ.
Адаптация облигатных метилотрофных бактерий М. flagellatum. В связи с важностью препаративного аспекта получения дейтерий-меченных соединений в рамках данной работы была изучена возможность адаптации различных штаммов-продуцентов БАС к росту на средах с максимальными концентрациями тяжелой воды (D2O). Для этого были проверены представители различных таксономических групп метилотрофных бактерий, имеющихся в коллекции ГосНИИ Генетики: L-лейцин-продуцирующий штамм облигатных метилотрофных бактерий М. flagellatum (ileu), реализующий 2-кего-3-дезокси-6-фосфогдюконат-альдолазный (КДФГ) вариант рибулё'зо-5- монофосфатного (РМФ) цикла ассимиляции углерода и L-фенилаланин-продуцирующий штамм факультативных метилотрофных бактерий В. methylicum (leu), ассимилирующий метанол по РМФ- циклу.
Для проведения адаптации был выбран ступенчатый режим увеличения концентрации тяжёлой воды в ростовых средах, так как мы предположили, что постепенное привыкание организма к тяжёлой воде будет оказывать благоприятный эффект на скорость роста культуры. При этом штамм М. flagellatum обнаружил повышенную чувствительность к тяжёлой воде: ингибирование роста бактерий наблюдалось при концентрациях D2О в среде 74,5 об.%. Роста бактерий на более высокой концентрации тяжёлой воды достичь не удалось. В связи с этим, в экспериментах по изучению уровней включения дейтерия в аминокислоты использовали препараты культуральной жидкости и биомассы М. flagellatum, полученные со среды, содержащей 74,5 об.% тяжёлой воды. Концентрация экзогенного дейтерометанола CD3OD составляла, как обычно, 1 об.%.
Адаптация факультативных метилотрофных бактерий В. methylicum. Попытки адаптировать штамм В. methylicum к росту при сохранении способности к биосинтезу L-фенилаланина на максимально дейтерированной среде привели к желаемому результату. К данному штамму метилотрофных бактерий был применён специально разработанный нами подход по адаптации, который заключался в серии из пяти адаптационных пассажей исходной культуры на агаризованных средах (с добавкой 2 об. % дейтерометанолом CD3OD) при ступенчатом увеличении концентраций экзогенной тяжёлой воды (от 0; 24,5; 49,0; 73,5 об% до 98 об% D2O) и последующей селекции устойчивых к тяжёлой воде клонов бактерий. При этом последовательно отбирали отдельные колонии, выросшие на средах, содержащих тяжёлую воду. Затем их пересевали на среды с большей степенью дейтерированпости, включая среду с 98 об.% тяжёлой водой (степень выживаемости бактерий на конечной полностью дейтерированной среде составляет не более 40%).
Полученный результат в опытах по адаптации В. methylicum к тяжёлой воде, позволил использовать гидролизаты его биомассы, а также саму биомассу, полученную в ходе многоступенчатой адаптации к D2O в качестве полноценных ростовых субстратов для выращивания бациллярных штаммов В. subtillis и В. amytoliquefaciens, а также штамма галофильных бактерий Н. halobium ET1001.
Адаптация бацилл В. subtittis и В. amyloliqucfaciens. В следующих опытах была исследована способность к росту на тяжёлой воде бациллярных штаммов В. subtillis (his, tyr, ade, иrа), и В. amyloliquefaciens (ade, иrа), продуцентов инозина и тимидина, соответственно. Мы предположили, что замедление роста бактерий на минимальных средах, содержащих тяжёлую воду могло быть обусловлено появлением ауксотрофности по отдельным ростовым факторам. Чтобы проверить это предположение, в дальнейшем мы использовали комплексные среды. Как и предполагалось, обе культуры удалось адаптировать к дейтерию путём рассева на твёрдые среды, приготовленные из 99,9 ат.% тяжёлой воды. Они сразу обнаружили нормальный рост на тяжёлой воде. У штаммов В. subtilis и В. amyloliquefaciens при росте на тяжёлой воде было отмечено сохранение высокого уровня продукции по инозину и тимидину (3,9 и 3,0 г/л соответственно).
Адаптация галофильных бактерий Н. halobium ET 1001. В случае с Н. halobium ET 1001 адаптацию проводили как на агаре, содержащим 99,9 ат.% тяжёлую воду путём рассева штамма до отдельных колоний, так и на жидкой среде с тяжёлой водой. В обычных для этой бактерии условиях культивирования (37°С, на свету) в клетках синтезировался фиолетовый пигмент по всем характеристикам не отличающийся от нативного бактериородопсина.
2. ИЗУЧЕНИЕ РОСТА И БИОСИНТЕЗА БАС ПОЛУЧЕННЫМИ ШТАММАМИ
Изучение ростовых характеристик М. jlagellatum на средах, содержащих CH3OH/CD3OD и D2O, а также 13СНзОН. Данные по росту штамма М. Jlagellatum на минимальных средах с добавкой 1 об.% метанола СНзОН и его меченных аналогов (СDзOD/13СНзОН) и содержащих ступенчато увеличивающиеся концентрации тяжёлой воды приведены в таблице I. Как видно из таблицы I, на средах, содержащих тяжёлую воду и изотопные аналоги метанола – дейтеро-метанол CD3OD и 13С-метанол 13CH3OH, выходы микробной биомассы составили 81% и 72% соответственно, а на средах с 74,5 об.% тяжёлой водой выход биомассы составил 29%, что в 3,4 раза ниже, чем в контрольных экспериментах, когда использовали обычную воду и метанол СНзОН (табл. 1, опыты 1,3,8). Как видно, способность к росту у М. flagellatum сохранялась лишь в среде, содержащей 74,5 об.% тяжёлой воды. Выше этой концентрации наблюдалось ингибирование скорости роста бактерий.
Таблица 1. Влияние изотопного состава среды на рост штамма M.flagelaum.
| Номер Компоненты среды, об% Величина Выход Время опыта лаг-фазы биомассы генерации Н2О D2O СНзОН СDзОD часы % ч | |||||||
| 1 | 99,0 | 0 | 1,0 | 0 | 0 | 100 | 1,1 |
| 2 | 99,0 | 0 | 0,5 | 0,5 | 0,2 | 91,0 | 0,8 |
| 3 | 99,0 | 0 | 0 | 1,0 | 0,8 | 81,0 | 1,0 |
| 4 | 49,5 | 49,5 | 1,0 | 0 | 2,4 | 76,0 | 1,4 |
| 5 | 49,5 | 49,5 | 0,5 | 0,5 | 5,7 | 75,0 | 1,2 |
| 6 | 49,5 | 49,5 | 0 | 1,0 | 6,7 | 70,0 | 1,3 |
| 7 | 24,5 | 74,5 | 1,0 | 0 | 5,6 | 29,0 | 1,4 |
| 8 | 99,0 | 0 | 1,0 'ЗСНзОН | 0 | 0,1 | 72,0 | 1,0 |
Как и следует из литературных данных (Складнее Д. А, 1990), введение стабильного изотопа углерода |3С не приводит к летальным последствиям для клетки, что мы и наблюдали в случае с М. flage/talum. В целом, полученные для М. flagellatum данные могут свидетельствовать о том, что адаптация к тяжёлой воде определяется как видовой специфичностью метилотрофных бактерий, так и особенностями их метаболизма. Кроме этого, из таблицы 1 следует, что данный подход можно эффективно использовать для введения в синтезируемые БАС двойной изотопной метки (дейтерий- и изотоп углерода 15C).
Изучение ростовых и биосинтетических характеристик Б. methylicum на средах, содержащих CH3OH/CD3OD и D2O. Данные по росту исходного и адаптированного к тяжёлой воде штамма В. methylicum и максимальному уровню накопления L-фенилаланина в культуральной жидкости на минимальных средах с добавкой 2 об.% метанола и его дейтерированного аналога СН3OН/CD3OD, содержащих ступенчато увеличивающиеся концентрации тяжёлой воды, представлены в таблице 2. Как видно из табл. 2, в отсутствии дейтерий-меченных субстратов продолжительность лаг-фазы не превышала 24 ч (см. табл. 2, опыт 1). С увеличением концентрации тяжёлой воды в среде продолжительность лаг-фазы увеличивалась до 64,4 ч на средах с 98 об.% тяжелой водой и 2 об.% CD3OD (табл. 2, опыт 10). Отмечено, что длительность времени клеточной генерации с увеличением степени изотопного насыщения среды дейтерием постепенно увеличивается, достигая 4,9 часов на максимально дейтерированной среде (табл. 2, опыт 10).
Таблица 2.
Влияние изотопного состава среды на рост штамма В. mehylicum и уровень накопления L-фенилаланина в культуральной жидкости*.
| Номер Компоненты среды, об% лаг-фаза Выход Время Выход опыта биомассы генер. L-Phe, Н2 О D2O СH3ОН CD3OD ч % ч % | ||||||||
| 1 | 98 | 0 | 2 | 0 | 24,0 | 100 | 2,2 | 100 |
| 2 | 98 | 0 | 0 | 2 | 30,3 | 92,3 | 2,4 | 99,1 |
| 3 | 73,5 | 24,5 | 2 | 0 | 32,1 | 90,6 | 2,4 | 96,3 |
| 4 | 73,5 | 24,5 | 0 | 2 | 34,7 | 85,9 | 2,6 | 97,1 |
| 5 | 49,0 | 49,0 | 2 | 0 | 40,5 | 70,1 | 3,0 | 98,0 |
| 6 | 49,0 | 49,0 | 0 | 2 | 44,2 | 60,5 | 3,2 | 98,8 |
| 7 | 24,5 | 73,5 | 2 | 0 | 45,8 | 56,4 | 3,5 | 90,4 |
| 8 | 24,5 | 73,5 | 0 | 2 | 49,0 | 47,2 | 3,8 | 87,6 |
| 9 | 0 | 98,0 | 2 | 0 | 60,5 | 32,9 | 4,4 | 79,5 |
| 10 | 0 | 98,0 | 0 | 2 | 64,4 | 30,1 | 4,9 | 71,5 |
| 10' | 0 | 98,0 | 0 | 2 | 39,9 | 87,2 | 2,9 | 95,0 |
'Данные (1-10) приведены для В. methylicum, не адаптированного к средам с высоким содержанием дейтерия.
Данные 10' приведены для адаптированного В. methylicum.
Как видно из табл. 2, опыт 2, дейтерометанол CD3OD не вызывал существенного ингибирования роста и не оказывал влияния на выход микробной биомассы, в то время как на средах с 98 об.% тяжёлой водой микробный рост подавлялся. Так, на среде, содержащей 98 об.% тяжёлой воды и 2 об.% дейтерометанола СDзОD, выход микробной биомассы был снижен в 3,3 раза no-сравнению с контролем. Важно то, что выход микробной биомассы и уровень накопления L-фенилаланина в культуральной жидкости при росте адаптированного к тяжёлой воде штамма В. inethylicum в полностью дейтерированной среде изменяются по сравнению с контрольными условиями на 12,8% и 5% соответственно (табл. 2, опыт 10').
За счёт использования данного штамма В. methylicum удалось выделить порядка 1 г L-Phe из 1 л среды.
Исследование биосинтеза L-фенилаланина штаммом В. methylicum. Общей особенностью биосинтеза L-Phe в протонированных средах было значительное увеличение его продукции на ранней фазе экспоненциального роста В. inethylicum, когда выход микробной биомассы был незначителен (рис. I).
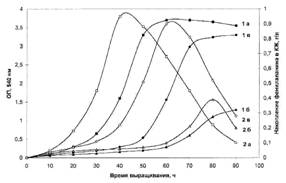
Рис. I. Динамики роста В. methylicum (la, 10'а, 10а) и накопления L-Phe в культуральной жидкости (16, 10'б, 106) на средах с различным изотопным составом: 1 а,б - исходный микроорганизм на протоннрованной среде М9; 10' а,б -адаптированный В. methylicum на полностью дейтерированной среде; 10 а,б - еадаптированный микроорганизм на полностью дейтерированной среде.
Во всех изотопных экспериментах наблюдалось ингибирование биосинтеза L-фенилаланина на поздней фазе экспоненциального роста и снижение его концентрации в ростовых средах. Согласно данным по микроскопическому исследованию растущей популяции микроорганизмов, наблюдаемый характер динамики секреции L-Phe не коррелировал с качественными изменениями ростовых характеристик культуры на различных стадиях роста, что служило подтверждением морфологической однородности микробной популяции. Скорее всего, накопленный в процессе роста фенилаланин ингибировал ферменты собственного пути биосинтеза. Кроме того, мы не исключаем возможность, что при ферментации без рН-статирования может происходить обратное превращение экзогенного фенилаланина в интермедиаторные соединения его биосинтеза, что отмечено в работах других авторов (Ворошилова Э. Б., Гусятипер М. М., 1989). Данные по исследованию культуральной жидкости методом тонкослойной хроматографии (ТСХ) показали, что кроме L-фенилаланина данный штамм В. methylicum синтезирует и накапливает в культуральной жидкости другие аминокислоты (аланин, валин, лейцин, изолейцин), четко детектируемые масс-спектрометрическим анализом (см. след, главу).
Изучение качественного и количественного состава внутриклеточных сахаров В. subtilis. В ходе выполнения работы был изучен качественный и количественный состав внутриклеточных Сахаров при росте В. subtilis на среде с 99,9 ат.% тяжёлой воды (см. табл. 3). Как видно из таблицы 3, в гидролизатах биомассы данного штамма фиксируются глюкоза, фруктоза, рамноза, арабиноза, сахароза и мальтоза.
Таблица 3.
Качественный и количественный состав внутриклеточных Сахаров В. subtilis при росте на 99,9 %тяжёлой воде.
| Компонент Содержание в биомассе, % Рост на Н2О Рост на 99,9% D2O | ||
| Глюкоза | 20,01 | 21,40 |
| Фруктоза | 6,12 | 6,82 |
| Рамноза | 2,91 | 3,47 |
| арабиноза | 3,26 | 3,69 |
| мальтоза | 15,30 | 11,62 |
| сахароза | 8,62 | - |
Изучение аминокислотного состава биомассы метилотрофных бактерий В. tnethylicum. Аминокислотный состав суммарных белков биомассы В. methylicum, полученного в ходе многоступенчатой адаптации к тяжёлой воде показан в таблице 4. Результаты исследования показали небольшое снижение содержания в дейтерированном белке Ala, Leu и Нis по сравнению с белком, полученным на обычной воде (табл. 4).
Таблица 4.
Качественный и количественный состав аминокислот общих белков биомассы В. methylicum.
| Аминокислота Содержание в белке, % Рост на Н2О Рост на 98% D2O | ||
| Gly | 8,03 | 9,69 |
| Ala | 12,95 | 13,98 |
| Val | 3,54 | 3,74 |
| Leu | 8,62 | 7,33 |
| His | 4,14 | 3,64 |
| Phe | 3,88 | 3,94 |
| Tyr | 1,56 | 1,82 |
| Asp | 7,88 | 9,59 |
| Glu | 11,68 | 10,38 |
| Lys | 4,37 | 3,98 |
| His | 3,43 | 3,72 |
| Thr | 4,81 | 5,51 |
| Met | 4,94 | 2,25 |
| Arg | 4,67 | 5,27 |
Изучение ростовых и биосинтетических характеристик В. subtilis на средах, содержащих тяжёлую воду и гидролизаты метилотрофных бактерий. Кривые, отражающие динамику роста, ассимиляции глюкозы и накопление инозина в культуральной жидкости штаммом В. subtilis в условиях протонированной среды и среды, с 99,9 ат.% тяжёлой воды представлены на рис. 2.
Как видно из рис. 2, при переносе клеток со стандартной на дейтерированную среду выход микробной биомассы, продолжительность лаг-фазы и длительность времени клеточной генерации в целом изменяются незначительно. При росте исходного штамма В. subtilis па среде, содержащей обычную воду уровень накопления инозина в культуральной жидкости достигал величины 17,3 г/л после пяти суток культивирования (рис. 2). Уровень накопления инозина на дейтерированной среде был снижен в 4,4 раза по-сравнению с исходным штаммом на протонированной среде (рис. 2). Низкие уровни секреции инозина на дейтерированной среде коррелируют со степенью конверсии глюкозы в этих условиях. Так, кривая конверсии глюкозы на полностью дейтерированной среде имела меньший угол наклона, чем на среде с обычной водой, что свидетельствует о том, что при росте на дейтерированной глюкоза расходуется менее эффективно (рис. 2).
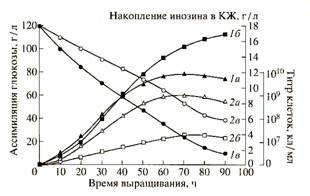
Рис.2. Динамики роста B.subtilis (1a, 2a), конверсии глюкозы (1б,2б) и накопления инозина в культуральной жидкости (1в,2в) на средах с различным изотопным составом: 1 а,б,в-B. Subtilis на обычной протонированной среде; 2 а,б,в-B.subtilis на полностью дейтерированной среде с гидролизатом дейтеро-биомассы метилотрофных бактерий.
Полученные для исследуемых микроорганизмов данные, в целом, подтверждают устойчивое представление о том, что адаптация клетки к тяжёлой воде является фенотипическим явлением, поскольку адаптированные к тяжёлой воде клетки возвращаются к нормальному росту и биосинтезу в протонированных средах после некоторого лаг-периода. В то же время обратимость роста на D2O/H2O-cpeдax теоретически не исключает возможности того, что этот признак стабильно сохраняется при росте в тяжёлой воде, но маскируется при переносе клеток на дейтерированную среду. Можно предположить, что клетка реализует лабильные адаптивные механизмы, которые способствуют функциональной реорганизации работы ферментных систем в тяжёлой воде. Также не исключено, что наблюдаемые при адаптации эффекты связаны с образованием в тяжёлой воде более прочных и стабильных связей, чем связей с участием водорода. По теории абсолютных скоростей разрыв С-Н-связей может происходить быстрее, чем C-D-связей, подвижность дейтерия D+ меньше, чем подвижность протия Н+, константа ионизации D2O в 5 раз меньше константы ионизации Н2О (Crespy J., Kalz H.H., 1979). С точки зрения физиологии, наиболее чувствительными к замене протия на дейтерий могут оказаться аппарат биосинтеза макромолекул и дыхательная цепь, т. е. именно те клеточные системы, которые используют высокую подвижность протонов и высокую скорость разрыва водородных связей.
3. ИЗУЧЕНИЕ СТЕПЕНЕЙ ВКЛЮЧЕНИЯ ИЗОТОПОВ ДЕЙТЕРИЯ и УГЛЕРОДА 13С в МОЛЕКУЛЫ ЭКЗОГЕННЫХ АМИНОКИСЛОТ B. methylicum и М. flagellatum.
Получение препаратов культуральных жидкостей, содержащих экзогенные дейтерий - и 13С-аминокислоты. Дейтерий-меченные аминокислоты были выделены в составе препаратов лиофилизованных интактных культуральных жидкостей, свободных от белков и полисахаридов, при росте штамма В. methylicum на минимальных средах с добавкой 2 об% метанола СНзОН и с различным содержанием тяжёлой воды. |3С-аминокислоты были получены за счет культивирования штамма М, flagellatum на среде, содержащей обычную воду и 1 об% 13С-метанол |3СНзОН. Данные по степеням включения дейтерия и 13С в молекулы экзогенных аминокислот двух исследуемых штаммов приведены в таблице 5. Во всех анализируемых образцах культуральной жидкости независимо от рода штаммов методом масс-спектрометрии электронного удара были обнаружены аланин, валин, лейцин/изолейцин и фенилаланин (табл. 5). В масс-спектрах дериватизованной культуральпой жидкости M.flagellatwn в дополнение к вышеобозначенным аминокислотам также фиксировался глицин.
Получение метиловых эфиров дансил-и карбобензокси-проазводных аминокислот. Степени включения изотопов дейтерия и мзотопа углерода 13С в мультикомпонептные смеси аминокислот в составе культуральной жидкости и белковых гидролизатов определяли методом высокочувствительной масс-спектромстрии электронного удара метиловых эфиров Dns-аминокислот или в виде Z-производных аминокислот после их препаративного разделения методом обращённо-фазовой высокоэффективной жидкостной хроматографии ОФ ВЭЖХ.
Аналитическое и препаративное разделение Z-производных аминокислот проводили методом ОФ ВЭЖХ, разработанным Егоровой Т. А. (Егорова Т. А., 1993). Степени хроматографической чистоты выделенных из культуральных жидкостей В. methylicwn и М. flagellatum 2-й Dns-производных дейтерий- и 13С-аминокислот составили 93-95%, а выходы 65-87% соответственно.
Предложенная нами модификация метода получения производных аминокислот заключалась в прямой химической обработке препаратов культуральной жидкости, полученной после отделения клеток, DnsCI (и ZCI) и CN2H2. Реакцию проводили в щелочной среде в водно-органическом растворителе в соотношении DnsCl (ZCI) -аминокислота, равным 5:1 (см. схему ниже).
Для лизина, гистидина, тирозина, серина, треонина и цистеина наряду с моно-производными было характерно образование ди-Z-(Dns)-производных: ди-Z,(Dns)-лизина, ди-Z,(Dns)-гистидина, О,N-ди-Z,(Dns5)-тирозина, О,N-ди-Z,(Dns)-серииа, O,N-AH-Z,(Dns)-Tpeoнина и N,S-ди-Z,(Dns)-цистеина (на схеме эти произодные не показаны). Кроме этого, из аргинина синтезировался три-Z,(Dns)-аргинин.
Летучесть Dns-и Z-производных аминокислот при масс-спектрометрическом анализе повышали за счет дополнительной дериватизации по карбоксильной группе (этерификации) диазометаном. Выбор диазометана в качестве этерифицирующего реагента был связан с необходимостью проведения реакции в мягких условиях, исключающих обратный изотопный (H-D-обмен в ароматических аминокислотах. При использовании диазометана происходило дополнительное N-метилирование по a-NH2-rpynne аминокислот, в результате чего в масс-спектрах метиловых производных аминокислот фиксировались дополнительные пики, соответствующие соединениям с молекулярной массой на 14 массовых единиц больше исходных.
Исследование степеней включения дейтерия в молекулу L-фенилаланина В. methylicum, полученного со сред с тяжёлой водой. Как видно из данных таблицы 2, рост данного штамма метилотрофных бактерий на средах с возрастающими концентрациями тяжёлой воды сопровождался снижением уровней накопления клеточной биомассы, увеличением времени генерации бактерий и продолжительности лаг-фазы при сохранении способности синтезировать и накапливать L-фенилаланин в ростовой среде. Поэтому было интересно изучить, как изменяются степени включения дейтерия в молекулу L-фенилаланина и других аминокислот В. methylicum в этих условиях.
Во всех опытах наблюдалось специфичное возрастание уровней изотопного включения дейтерия в молекулы аминокислот при ступенчатом увеличении концентраций тяжёлой воды в ростовой среде (табл. 5). Так, для индивидуальных аминокислот культуральной жидкости В. melhylicum, количество включённых атомов дейтерия по скелету молекул варьирует в пределах 49%-ной концентрации D2O и составляет для Phe 27,5%, Ala - 37,5%, Val - 46,3%, Leu/Ile - 47% (табл. 5). Аналогичное увеличение молекулярной массы аминокислот в зависимости от концентрации тяжёлой воды в среде было зафиксировано во всех экспериментах.
Таблица 5.
Степени включения дейтерия- и изотопа углерода 13С в молекулы секретируемых аминокислот В. melhylicum* и M. flagellation**.
| Аминокислоты | Содержание :Н2О в среде, об% 24,5 49,0 73,5 98,0 | 13СН3ОН 1 % | |||
| Gly | - | • | - | - | 60,0 |
| Ala | 24,0 | 37,5 | 62,5 | 77,5 | 35,0 |
| Val | 20,0 | 46,3 | 43,8 | 58,8 | 50,0 |
| Leu/Ile | 15,0 | 47,0 | 46,0 | 51,0 | 38,0 |
| Phe | 15,0 | 27,5 | 51,3 | 75,0 | 95,0 |
* Данные по включению дейтерия в аминокислоты приведены для В. methyticum при росте на средах, содержащих 2 об.% СН3ОН и 24,5; 49,5; 73,5; 98,0 об.% D2O. "Данные по включению 13С приведены для М. flagellatum при росте на среде, содержащей 1 об.% 13СН3ОН и 99 об.% Н2О.
Исследование степеней включения дейтерия в сопутствующие аминокислоты В. meihylicum на средах с тяжёлой водой. В масс-спектрах всех исследуемых образцов культуральной жидкости Б. melhylicum кроме основной секретируемой аминокислоты (фенилаланин) были обнаружены примеси, метаболически с ней связанных аланина, валина и лейцина/изолейцина {на уровне 3-5 мМ). В опыте, где концентрация тяжёлой воды в среде составила 49 об.% (таблица 5, опыт 5), изотопный состав фснилаланина характеризовался увеличением молекулярной массы на 4,1 единицу, аланина на 2,5 единицы, валина - 3,5 единицы, а лейцина/изолейцина - на 4,6 единиц. Таким образом, в отличии от фенилаланина, количество включенного дейтерия в последних трех аминокислотах сохраняет стабильное постоянство в довольно широком интервале концентраций экзогенной тяжёлой воды (от 49 об.% до 98 об.%).
В связи с тем, что штамм - продуцент фенилалапина В. methylicum был ауксотрофом по лейцину, эту аминокислоту в немеченном виде добавляли в ростовую среду, содержащую 98 об.% тяжёлой воды. Как показали наши исследования по включению дейтерия к молекулы экзогенных аминокислот, в условиях ауксотрофности по лейцину степень изотопного обогащения лейцина, а также метаболически связанных с ним аминокислот немного ниже, чем для других аминокислот. Так, при росте В. methylicum на среде, содержащей 98 об.% тяжёлой воды и немеченный L-Leu, степени включения дейтерия в Leu составили 51,0%, Ala -77,5%, Val - 58,8% (табл. 5). Суммируя полученные данные, можно сделать вывод о сохранении минорных путей метаболизма, связанных с биосинтезом лейцина de novo. Другим логическим объяснением наблюдаемого эффекта может быть ассимиляция клеткой немеченного лейцина из среды на фоне биосинтеза меченного изолейцина de novo.
Исследование степеней включения дейтерия в L-Phe В. methylicum в максимально дейтерированной среде. Мы предположили, что за счёт ауксотрофности штамма В. methylicum по лейцину, уровни включения дейтерия в секретируемыи фенилаланин на фоне максимальных концентраций тяжёлой воды могут быть ниже теоретически допустимых вследствие функционирования в клетке ряда биохимических реакций, связанных с ассимиляцией протонированного лейцина извне. Как мы и ожидали, отмеченная особенность лучше всего проявлялась при биосинтезе фенилаланина на дейтерированной среде, в которой единственным протонированным соединением, кроме метанола, являлся лейцин (см. табл. 5, опыт 9). В этом опыте степень дейтерированности L-фенилаланина составила 75%, т.е. только шесть атомов (из восьми в углеродном скелете) в молекуле" фенилаланина биосинтетически замещены на дейтерий. Согласно данным масс-спектрометрического анализа, атомы дейтерия распределены по положениям С1-С6 ароматической части фенилаланина и сопредельному положению b, причем, как миниум четыре из них могут быть локализованы в самом бензольном кольце молекулы фенилаланина. Результат по получению L-Phe с данным характером включения метки очень важен для биотехнологического использования и имеет существенные преимущества по-сравнению с химическим (Н-D)-обменом (Griffiths D. V., 1986).
Исследование степени включения дейтерия в молекулу фенилаланина за счёт конверсии дейтерометанола CD3OD в В. methylicum.
Контроль за включением дейтерия в молекулу L-Phe за счет конверсии дейтерометанола CD3OD при росте бактерий на среде, содержащей обычную воду и 2 об.% дейтерометанол CD3OD (соответствуют опыту 2, табл. 1) показал незначительное количество дейтерия, которое поступает в молекулу L-фенилаланина вместе с углеродом CD3OD, Процент дейтерирования фенилаланина был вычислен по величине пика с m/z 413 за вычетом вклада пика примеси природного изотопа (не более 4%). Полученный результат может быть объяснён разбавлением дейтериевой метки за счёт протекания как биохимических процессов, связанных с распадом дейтерометанола CD3OD при его ассимиляции клеткой, так и реакциями изотопного обмена и диссоциации в тяжёлой воде. Так, из четырёх атомов дейтерия, имеющихся в молекуле СDзOD, лишь один атом дейтерия при гидроксильной группе -OD самый подвижный и поэтому легко диссоциирует в водной среде с образованием СDзОН. Три оставшихся атома дейтерия в составе СDзОН входят в цикл ферментативного окисления метанола, который, в свою очередь, мог привести к потере дейтериевой метки за счёт образования соединений более окисленных, чем метанол. В частности, такое включение дейтерия в молекулу L-фенилаланина подтверждает классическую схему ферментативного окисления метанола до формальдегида в клетках метилотрофов, который лишь после этого ассимилируется у данного штамма метилотрофных бактерий РМФ-путем фиксации углерода (Nesvera J., 1991).
Исследование степеней включения изотопа углерода 13С в молекулы экзогенных аминокислот М. flagellatum за счёт биоконверсии 13СНзОН.
Наши исследования подтвердили, что для получения |3С-аминокислот за счет микробной конверсии 13СНзОН предварительная адаптация не является лимитирующим этапом, поскольку этот., субстрат не оказывает негативного биостатического эффекта на ростовые и биосинтетические характеристики метилотрофов. При росте М. flagellatum на среде, содержащей 99 об.% воду и 1 об.% |3СНзОН клетка продуцирует лейцин, a также глицин, фланин, валин и фенилаланин. Как видно из таблицы 5, уровни изотопного включения |3С в Gly, Ala, Val и Phe составляют 60, 35, 50 и 95% соответственно. При этом низкая степень включения изотопа углерода |3С в метаболически связанные с изолейцином аминокислоты обусловлена эффектом ауксотрофиости бактерий в изолейцине, который добавляли в ростовую среду в немеченном виде.
4. ИЗУЧЕНИЕ СТЕПЕНЕЙ ВКЛЮЧЕНИЯ ИЗОТОПОВ ДЕЙТЕРИЯ и УГЛЕРОДА 13С в АМИНОКИСЛОТНЫЕ ОСТАТКИ СУММАРНЫХ БЕЛКОВ M. methylkiim и М.flagellatum.
Выделение дейтерий- и 1}С-алшнокислот из белковых гидролизатов. Поскольку при работе с микробной биомассой возникают проблемы, связанные с очисткой от сопутствующих компонентов, было необходимо применять специальные подходы при выделении фракции суммарных белков из бактериальных источников.
При выделении фракции суммарных белков биомассы метилотрофных бактерий (В. methylicum, M. flagellatum) учитывалось наличие в них углеводов. Мы использовали богатые по белку штаммы бактерий со сравнительно небольшим содержанием углеводов в них, гидролизу в качестве фракции суммарных белков подвергали остаток после исчерпывающего отделения пигментов и липидов экстракцией органическими растворителями (метанол-хлороформ-ацетон}.
Во всех случаях гидролиз белков проводили в 6 н. растворе DC1 (3 масс.% фенола в D2O) или в 4 н. растворе Ва(ОН):для предотвращения реакций обратного изотопного обмена (H-D) в ароматических аминокислотах и их разрушения.
Дейтерий- и 13С-меченные аминокислоты в составе гидролизатов суммарного белка биомассы были разделены методом ОФ ВЭЖХ со степенью хроматографической чистоты 93-96% и выходами 75-89% в условиях, аналогичных таковым для разделения секретируемых аминокислот (табл. 6). Хотя в таблице 6 приведены данные только для 10 аминокислот, очевидно, что в остальных аминокислотах уровни изотопного включения сопоставимы, хотя они не детектируются данным методом. Это предположение подтверждается данными по разделению белковых гидролизатов метилотрофных бактерий методом ионнообменной хроматографии на колонке "Biotronic LC 5001", где детектируется уже 15 аминокислот (см. рис. 4).
Исследование степеней включения дейтерия в аминокислотные остатки белка В. methylicum па средах с тяжёлой водой.
Общие принципы изучения степени изотопного обогащения молекул аминокислот при данном способе введения метки продемонстрированы на примере анализа включения дейтерия в мультикомпонентные смеси аминокислот, полученные после гидролиза суммарных белков биомассы в 6 н. DCl и 4 н. Ва(ОН).
Во всех -экспериментах по научению содержания дейтерия в аминокислотных остатках белка наблюдалась корреляция между степенью изотопного насыщения среды и уровнями включения дейтерия в аминокислоты (табл. 6), Например, для индивидуальных аминокислот белковых гидролизатов количество включенных атомов дейтерия по скелету молекулы варьирует незначительно в пределах 49%-ной концентрации тяжёлой воды и составляет для Ala 45%, Val - 36,3%, Leu/Ile - 42%, Phe -37,5%.
Таблица 6.
Степени включения D и 13С в аминокислотные остатки общих белков биомассы В. melhyiicum* и М. flagellatum**.
| Аминокислоты | Содержание D2O в среде, об% 24,5 49,5 73,5 98,0 | 13СНзОН 1 об% | |||
| Gly | 15,0 | 35,0 | 50,0 | 90,0 | 90,0 |
| Ala | 20,0 | 45,0 | 62,5 | 97,5 | 95,0 |
| Val | 15,0 | 36,3 | 50,0 | 50,0 | 50,0 |
| Leu/lie | 10,0 | 42,0 | 45,0 | 49,0 | 49,0 |
| Phe | 24,5 | 37,5 | 50,0 | 95,0 | 80,5 |
| Туr | 20,0 | 48,8 | 68,8 | 92,8 | 53,5 |
| Ser | 15,0 | 36,7 | 47,6 | 86,6 | 73,3 |
| Asp | 20,0 | 36,7 | 60,0 | 66,6 | 33,3 |
| Glu | 20,0 | 40,0 | 53,4 | 70,0 | 40,0 |
| Lys | 10,0 | 21,1 | 40,0 | 58,9 | 54,4 |
'Данные по включению дейтерия в аминокислоты приведены для В. melhyiicum при росте на средах, содержащих 2 об.% СН3ОН и 24,5; 49,5; 73,5; 98,0 об.% D2O. **Данные по включению 13С приведены для М. flagellatum при росте на среде, содержащей 1 об.% 13СН3ОН и 99 об.% Н2О.
Исследование степеней включения дейтерия в аминокислотные остатки белка В. melhyiicum на максимально дейтерированной среде. Полученные данные свидетельствуют о возможности достижения максимальных уровней включения дейтерия в аминокислотные остатки белков за счет адаптации культуры В. melhyiicum к росту и биосинтезу на среде с максимальной концентрацией тяжёлой воды. Как видно из таблицы 6, при росте В. methylicum на среде, содержащей 98 об.% тяжёлой воды, степени включения дейтерия в остатки Gly, Ala и Phe составляют 90, 97,5 и 95%, т.е. уровень мечения можно считать униформным. Низкие степени включения дейтерия в молекулы лейцина (49%), а также в метаболически связанных аминокислотах в этих условиях могут быть объяснены за счет ауксотрофности штамма в лейцине, который добавляли в среду культивирования в протонированной форме. Полученный результат по разбавлению дейтериевой метки в лейцине может быть объяснён сохранением доли минорных реакций в биосинтезе лейцина de novo.
Исследование степеней включения изотопа углерода I3C в аминокислотные остатки белка М.
flagellation за счёт биоконверсии 13СH3ОН.
В экспериментах по включению изотопа углерода |3С в молекулы белков за счёт биоконверсии 13СH3ОН метилотрофными бактериями М.flagellalum была показана эффективность мечения аминокислот изотопом углерода 13С. Так, в Phe детектировалось 80,5 % метки, в Ala - 95 %, в Gly - 90% (см. табл. 6).
Во всех экспериментах степени включения дейтерия и изотопа углерода 13С в метаболически связанных аминокислотах обнаружили определённую коррелляцию. Так, степени изотопного обогащения валина и лейцина (семейство пирувата), фснилаланина и тирозина (семейство ароматических аминокислот} совпадают (табл. 6). Степени изотопного включения глицина и серина (семейство серина), аспарагиповой кислоте и лизина (семейство аспарагина) также имеют близкие величины. Сравнивая данные таблицы 5 и 6, можно заключить, что степени изотопного обогащения экзогенных аминокислот и соответствующих аминокислотных остатков суммарного белка, в целом, также коррелируют.
Как и в случае с экзогенными аминокислотами, низкие степени включения изотопа углерода 13С в остатки Leu при росте на 1 об.% 13СНзОН обусловлены ауксотрофностью бактерий в этой аминокислоте.
Таким образом, нам удалось достичь максимальных уровней включения стабильных изотопов в суммарные белки биомассы метилотрофных бактерий. Именно поэтому мы посчитали возможным использовать гидролизаты их биомассы для биосинтеза других изотопно - меченных БАС.
5. ИССЛЕДОВАНИЕ ВОЗМОЖНОСТИ ИСПОЛЬЗОВАНИЯ ГИДРОЛИЗАТОВ БИОМАССЫ МЕТИЛОТРОФНЫХ БАКТЕРИЙ В. methylkum в КАЧЕСТВЕ СУБСТРАТОВ ДЛЯ ПОЛУЧЕНИЯ [1',3',4',2,8-D5]-ИНОЗИНА.
Получение [1',3',4',2,8-D5]- инозина. В следующих экспериментах было апробировано использование дейтеро-компонентов биомассы метилотрофных бактерий, полученных в условиях многоступенчатой адаптации к тяжёлой воде для синтеза высокодейтерированных нуклеозидов (на примере инозина). [1,3',4',2,8-D5]-инозин был получен биосинтетически за счёт использования штамма-продуцента В. subtilis и выделен из культуральной жидкости по методике, включающей адсорбцию инозина на активированном угле, десорбцию спиртово-аммиачным раствором и перекристаллизацию из метанола. ТСХ инозина, с детекцией при 249 нм показала наличие в анализируемом образце единственного пятна с Rf = 0,55, -соответствующего по подвижности чистому инозину.
Особенности разработанного метода получения [1,3',4',2,8-D5]-инозина заключаются в следующих аспектах:
1. В способности высокоактивного штамма В. subtilis к росту и биосинтезу инозина на средах, содержащих максимальные концентрации тяжёлой воды;
2. Замене глюкозы и аминокислот, необходимых для роста этого штамма-ауксотрофа на гидролизаты дейтеро-биомассы В. methylicutn. При последующих ферментациях в качестве источника ростовых факторов можно использовать ту же дсйтеро-биомассу метилотрофных бактерий, либо биомассу самого штамма-продуцента, содержащую в своем составе соединения, которые могут служить источниками углерода и ростовых факторов;
3. В практически полном отсутствии отходов: согласно схеме, дейтеро-биомасса базового штамма, после гидролиза в 6 н. DC1 возвращается в цикл в качестве ростовых факторов;
4. В высокой степени изотопного обогащения дейтерий-мсченного инозина (62,5% атомов водорода в молекуле замещены на дейтерий);
5. В высоких выходах (3,9 г/л) меченного продукта.
Исследование уровня дейтерированности инозина. Места локализации дейтерия в молекуле инозина, были исследованы с помощью масс-спектрометрии FAB и спектроскопии ПМР (см. схему).
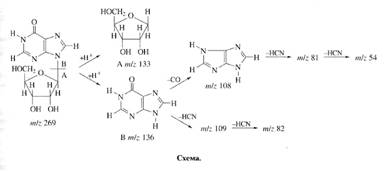
При анализе степени дейтерированности инозина учитывались следующие аспекты. Во-первых, вследствие того, что протоны в C1-C's положениях рибозной части молекулы инозина могли происходить из глюкозы, мы предположили,что характер биосинтетического включения дейтерия в рибозную часть молекулы инозина определяется, в основном, функционированием ряда процессов гексозо-моно-фосфатного (ГМФ) шунта, связанных непосредственно с ассимиляцией глюкозы и других сахаров. Во-вторых, многочисленные обменные процессы и внутримолекулярные перегруппировки, происходящие с участием тяжёлой воды могли также привести к специфическому включению метки по определенным позициям в молекуле инозина. Такими доступными позициями в молекуле инозина признаны, прежде всего, гидроксильные протоны -ОН и протоны при гетероатомах -NH (последние могут обмениваться на дейтерий в тяжёлой воде за счет кето-енольной таутомерии). Три атома дейтерия в рибозном остатке молекулы инозина могли происходить за счет функционирования многочисленных реакций ГМФ-шунта, два атома дейтерия а гипоксантинс также могли синтезироваться de novo (схема).
0 комментариев