Дихотомический путь
Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
Глиоксилатный цикл. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Навигация
Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
Механизмы дыхания растений
89666
знаков
0
таблиц
22
изображения
4.2 Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
В клетках растений наряду с гликолизом и циклом Кребса, являющимся главным поставщиком свободной энергии в процессах дыхания, существует и другой важнейший способ катаболизма гексоз — пентоэофосфатный путь (ПФП), в котором участвуют пятиуглеродные сахара (пентозы). Этот путь дыхания известен также как гексозомонофосфатный цикл, пентозный шунт или апотомическое окисление. Окисление глюкозы (глюкозо-6-фосфата) по этому пути связано с отщеплением первого (альдегидного) атома углерода в виде С02 (отсюда и название — апотомический путь).
Пентоэофосфатный путь дыхания открыт в 1935—1938 гг. в результате исследований О. Варбурга, Ф. Диккенса, В. А. Эн- гельгардта и позднее Ф. Липмана. Установлено, что все реакции ПФП протекают в растворимой части цитоплазмы клетс а также в пропластидах и хлоропластах. ПФП дыхания особенно активен в тех клетках и тканях растений, в которых интенсивно идут синтетические процессы, такие, как синтез липидных компонентов мембран, нуклеиновых кислот, клеточных стенок, фенольных соединений.
В ПФП АТР используется только для образования исходного продукта. ПФП, как и цикл Кребса, — циклический процесс, поскольку окисление глюкозы сопровождается регенерацией исходного субстрата ПФП — глюкозо-6-фосфата.
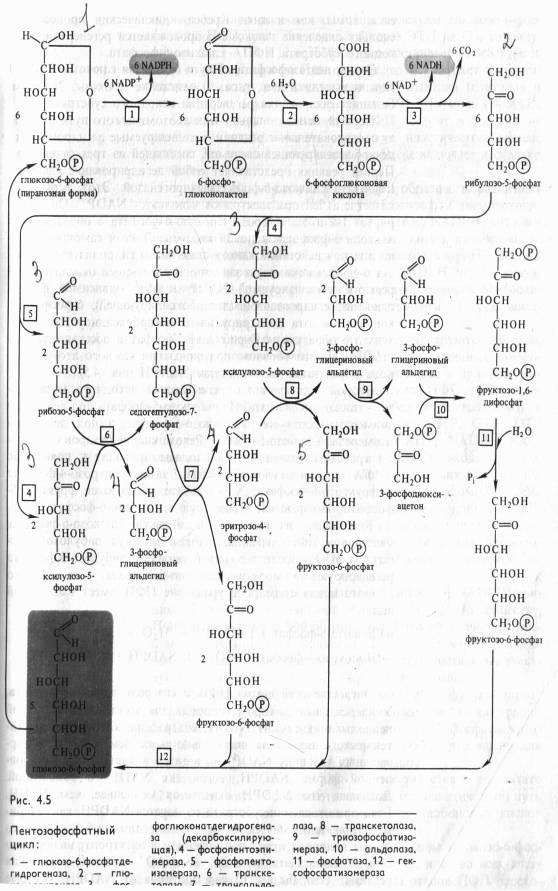
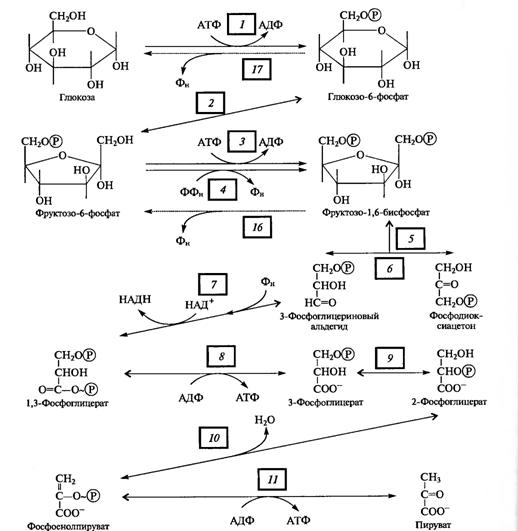
Этапы пентозофосфатного пути окислении глюкозы. В ПФП можно выделить два этапа: 1) окисление глюкозы, 2) рекомбинацию Сахаров для регенерации исходного субстрата.
Первый, окислительный, этап апотомического пути включает последовательные реакции, катализируемые дегидрогеназнодекарбоксилрующей системой, состоящей из трех ферментов. Первая реакция представляет собой Дегидрирование глюкозо- 6-фосфата глюкозо-6-фосфатдегидрогеназой. Этот фермент в качестве акцептора электронов использует NADP + . Он дегидрирует 1-й атом углерода глюкозо-6-фосфата с образованием лактона 6-фосфоглюконовой кислоты. Лактон самопроизвольно или под действием глюконолактоназы гидролизуется, образуя 6-фосфоглюконовую кислоту. В следующей окислительной реакции, катализируемой NADP- и Мп2 +-зависимой фосфо- глюконатдегидрогеназой (декарбоксилирующей), 6-фосфоглюконовая кислота дегидрируется и декарбоксилируется. В результате образуются О-рибулозо-5-фосфат и восстановленный NADPH. Таким образом, при окислении каждого атома углерода образуются две молекулы NADPH (рис. 4.5).
Второй этап связан с регенерацией исходного метаболита — глюкозо-6-фосфата. Из рибулозо-5-фосфата под действием эпимеразы образуется ксилулозо-5-фосфат, а под действием изомеразы — рибозо-5-фосфат. Рекомбинации Сахаров с участием транскетолазы и трансальдолазы приводят к появлению 3-ФГА и седогептулозо-7-фосфата, затем эритрозо-4-фосфата и фруктозо-6-фосфата; в результате образуются фруктозо-6- фосфаты, которые изомеризуются в глюкозо-6-фосфат.
Как видно из рис. 4.5, 6 молекул глюкозо-6-фосфата, участвуя в ПФП дыхания, дают 6 молекул рибулозо-5-фосфата и 6С02, после чего из 6 молекул рибулозо-5-фосфата регенерируют 5 молекул глюкозо-6-фосфата. Для каждого оборота цикла суммарное уравнение ПФП имеет следующий вид:
6 Глюкозо-6-фосфат + 12NADP + + 7Н20->- - 5Глюкозо-6-фосфат + 6С02 + 12NADPH + 12Н+ + Н3Р04
Энергетический выход ПФП и его роль в обмене веществ.
Универсальным донором водорода для электронтранспортной цепи дыхания служит NADH, содержание которого в растительных тканях всегда значительно выше, чем NADPH. В нормальных условиях NADP+ находится в клетках в восстановленной форме NADPH, тогда как NAD+ — в окисленной. Доказано, что NADPH окисляется медленнее, чем NADH. Если при окислении субстрата образуется NADPH, как, например, при апотомическом окислении глюкозо-6-фосфата, то атомы водорода перед поступлением в электронтранспортную цепь должны быть переданы на. NAD+ (трансгидрогеназцая реакция). Если бы все 12 пар протонов от NADPH, которые образуются при полном окислении молекулы глюкозо-6-фосфата по ПФП, были бы переданы через ЭТЦ на 02, то получилось бы 3 АТР х 12 = 36 АТР, что составляет 41,868 кДж х х 36 = 1507 кДж/моль. Практически это не уступает энергетическому выходу дихотомического пути дыхания (гликолиз и цикл Кребса), в котором образуется 1591 кДж/моль (38 АТР,)1
Однако основное назначение ПФП состоит в участии не столько в энергетическом, сколько в пластическом обмене клеток. Это участие в пластическом обмене включает несколько аспектов:
1. NADPH используется главным образом в различных синтетических реакциях.
2.В ходе пентозофосфатного цикла синтезируются пентозы, входящие в состав нуклеиновых кислот и различных нуклеотидов (пиридиновых, флавиновых, адениловых и др.). Для животных и других гетеротрофных организмов ПФП — единственный способ образования пентоз (рибоз и дезоксирибоз) в клетке. Рибозы необходимы для синтеза АТР, GTP, UTP и других нуклеотидов. Коферменты NAD+, NADP+, FAD, коэнзим А — тоже нуклеотиды и в их состав входит рибоза.
3.ПФП имеет большое значение как источник образования углеводов с различным числом углеродных атомов в цепи (от С3 до Су). Эритрозо-4-фосфат, возникающий в ПФП, необходим для синтеза шикимовой кислоты — предшественника многих ароматических соединений, таких, как ароматические аминокислоты, витамины, дубильные и ростовые вещества, лигнин клеточных стенок и др.
4.Компоненты ПФП (рибулозо-1,5-дифосфат, NADPH) принимают участие в темновой фиксации С02. По существу, ПФП представляет собой обращенный фотосинтетический (восстановительный) цикл Кальвина. Только две из 15 реакций цикла Кальвина специфичны для фотосинтеза, остальные участвуют в окислительном ПФП дыхания и гликолизе.
5.В хлоропластах окислительный ПФП функционирует в темноте, предотвращая резкое изменение концентрации NADPH в отсутствие света. Кроме того, триозофосфаты этого цикла в хлоропластах превращаются в 3-ФГК, что важно для поддержания в них уровня АТР в темноте.
Окисление глюкозы по ПФП осуществляется в результате 12 реакций, тогда как в дихотомический (гликолитический) путь через ПВК и далее через цикл ди- и трикарбоновых кислот включаются более 30 различных реакций.
Не следует, однако, думать, что окисление глюкозо-6-фосфата по схеме, представленной на рис. 4.5, идет во всех клетках до конца. Очень часто на одном из этапов ПФП перехо дит в гликолитический. Таким этапом может быть, в частности, транскетолазная реакция (рис. 4.5, реакция 8), в результате которой ксилулозо-5-фосфат и эритрозо-4-фосфат превращаются во фруктозо-6-фосфат и 3-ФГА — субстраты, общие для гликолиза и ПФП.
Похожие работы
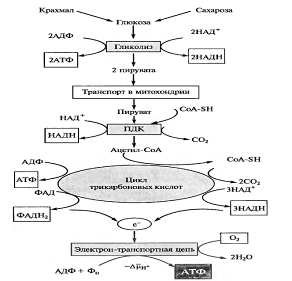
... ; обнаружены все ферменты, катализирующие превращение этих кислот; показано, что малонат — ингибитор сункцинатдегидрогеназы — тормозит окисление пирувата и резко снижает поглощение 02 в процессах дыхания у растений. Большинство ферментов цикла Кребса локализовано в мАТФиксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрии. Энергетический выход цикла Кребса, его ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... цитохромоксидаза, подтверждением чего служит также максимальная величина подавления дыхания азидом . Соотношения указанного типа не являются, однако, общей закономерностью, поскольку в ряде случаев максимальная интенсивность дыхания обнаруживается в клетках зоны растяжения. Такая закономерность выявляется при пересчете на белок, а также на единицу кислотонерастворимого фосфора. Вполне вероятно, ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев