Дихотомический путь
Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
Глиоксилатный цикл. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Навигация
Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Механизмы дыхания растений
89666
знаков
0
таблиц
22
изображения
8. Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Соссюр, работая с зелеными растениями в темноте, обнаружил, что они выделяют С02 даже в бескислородной среде. Л. Пастер нашел, что в темноте в отсутствие кислорода в растительных тканях наряду с выделением С02 образуется спирт, т. е. идет спиртовое брожение. Он пришел к выводу, что в растительных тканях, так же как и у бактерий, возможно спиртовое брожение.
Немецкий физиолог Э. Ф. Пфлюгер (1875), изучая дыхание животных объектов, показал, что лягушки, помещенные в среду без кислорода, некоторое время остаются живыми и при этом выделяют С02. Пфлюгер назвал это дыхание интрамолекулярным, т. е. дыханием за счет внутримолекулярного окисления субстрата. Предполагалось, что интрамолекулярное дыхание — начальный этап нормального аэробного дыхания. Эту точку зрения поддержал Б. Пфеффер — немецкий физиолог растений, который распространил ее на растительные организмы. На основе этих работ Пфеффером и Пфлюгером были предложены следующие два уравнения, описывающие механизм дыхания:
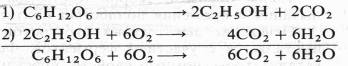
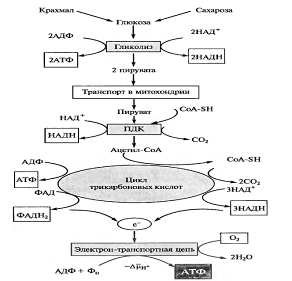
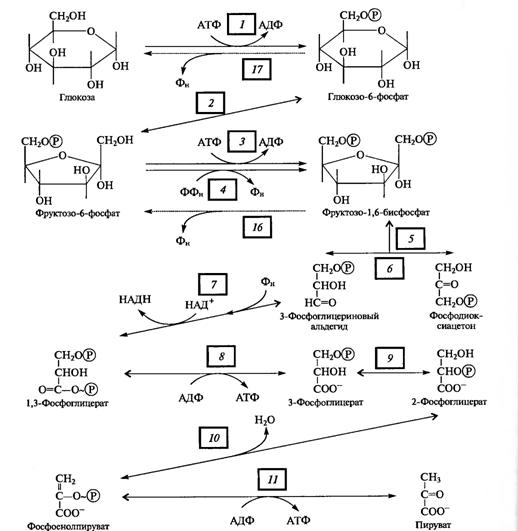
На первом, анаэробном, этапе происходит спиртовое брожение, образуются две молекулы этанола и две молекулы С02. Затем в присутствии кислорода спирт, взаимодействуя с ним, окисляется до С02 и Н20.
С. П. Костычев (1910) пришел к выводу, что это уравнение не соответствует действительности. Он экспериментально доказал, что этанол не может быть промежуточным продуктом нормального аэробного дыхания у растений по двум причинам: во-первых, он ядовит для растений и не может накапливаться, во-вторых, этанол окисляется растительными тканями значительно хуже, чем глюкоза. Костычев предложил свою формулу связи анаэробной и аэробной частей дыхания и различных видов брожения.
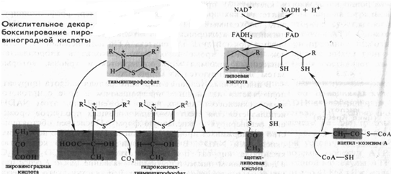
В опытах Костычева и его сотрудников (1912—1928) было показано, что если растительные ткани кратковременно выдержать в бескислородной среде, а затем дать кислород, то наблюдается резкое усиление дыхания, т.е. в ходе анаэробной фазы накапливаются промежуточные продукты, которые в присутствии кислорода быстро используются. Ингибиторы, блокирующие брожение, например NaF, блокируют и аэробное дыхание. Ингибиторный анализ (применение ингибиторов специфического действия), выделение и идентификация продуктов окислительного распада глюкозы привели Костычева к выводу о том, что промежуточным продуктом может быть уксусный альдегид. Благодаря работам немецкого биохимика К. Нейберга, Костычева и других стало очевидным, что дыхание и все виды брожения связаны между собой через пировиноградную кислоту (ПВК):
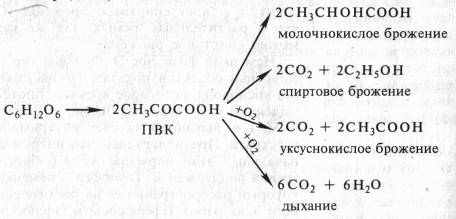
Таким образом, теория Костычева о генетической связи дыхания и брожения полностью подтвердилась.
Взаимосвязь дыхания с другими процессами обмена.
Дыхание тесно связано с другими процессами метаболизма. Необходимо подчеркнуть, что, несмотря на противоположную направленность двух центральных процессов растительного организма —.фотосинтеза и дыхания и на их рассредо- точенность в разных органеллах клетки, между ними существует тесная взаимосвязь. Прежде всего для протекания процесса дыхания органические вещества (субстраты).Таким субстратами в первую очередь являются углеводы, которые у зеленых растений образуются в процессе фотосинтеза. Вместе с тем превращение веществ в процессе фотосинтеза и дыхания идет через ряд сходных промежуточных продуктов. Особенно много сходного в превращениях между фотосинтетическим циклом Кальвина и реакциями пентозофосфатного пути дыхательного обмена. Как в том, так и в другом случае происходят взаимны превращения Сахаров с разной длиной углеродной цепочки (3, 4, 5, 6 и 7 углеродных атомов). По-видимому, несмотря на различное распределение (компартментацию) этих метаболитов в клетке, между ними существует обмен. Иначе го вор я, промежуточные. продукты дыхания могут быть использованы в процессе фотосинтеза. Одновременно возможен и обратный процесс. Много общего в энергетике фотосинтеза и дыхания в процессах фотосинтетического и окислительного фосфорилирования. Между этими двумя процессами возможен обмен энергетическими эквивалентами. АТФ, образовавшаяся на свету при фотосинтетическом фосфорилировании, может служить основным источником энергии для различных биосинтетических процессов, заменяя АТФ, образовавшуюся в процессе дыхания. С другой стороны, АТФ и НАДФ-Н, образовавшиеся в процессе дыхания, могут быть использованы для реакций цикла Кальвина. Имеются наблюдения, что на свету основными органеллами, поставляющими АТФ, являются хлоропласты.
Многие промежуточные продукты процесса дыхания являются основой биосинтеза важнейших соединений. Уже на протяжении первой, анаэробной фазы дыхания (гликолиз) триозофосфат, преобразуясь в глицерин, может служить источником для синтеза жиров. Пировиноградная кислота путем аминирования может дать аланин. Не менее важное значение имеют и промежуточные продукты цикла Кребса. Например, а-кетоглютаровая и щавелевоуксусная кислоты в процессе аминирования дают аминокислоты — глутаминовую и аспарагиновую. Благодаря реакции переаминиро- вания эти кислоты могут быть источником аминогруппы для других аминокислот и, таким образом, являться важнейшими промежуточными продуктами для синтеза как белка, так и пуриновых и пиримидиновых азотистых оснований. Янтарная кислота, образовавшаяся в цикле Кребса, дает основу для образования порфиринового ядра хлорофилла. Ацетил-КоА служит основой для образования жирных кислот. Поскольку имеется ряд реакций и процессов, благодаря которым отдельные компоненты извлекаются из цикла Кребса, должны быть и обратные процессы, поставляющие их в цикл. Если бы этого не было, скорость превращения в аэробной фазе дыхания заметно бы снизилось. Такими реакциями является окислительное дезаминирование аминокислот, приводящее к образованию органических кислот. Имеет значение также реакция карбоксилирования пировиноградной кислоты или ее фосфорилированой формы, в результате чего образуется щавелевоуксусная кислота. Основной процесс, при котором образуются пентозы в растении,— то пентозофосфатный путь дыхательного обмена. Пентозы входят в состав нуклеотидов, нуклеиновых кислот и ряда коферментов, в том числе таких важных, как никотинамидные (НАД и НАДФ), флавиновые (ФМН, ФАД). Пентозофосфатный путь дыхания является также источником образования эритрозо-4-фосфата. Эритрозофосфат, взаимодействуя с фосфоенолпируватом, образует шикимовую кислоту. Шикимовая кислота — материал для образования ряда ароматических аминокислот, например триптофана, а из трипсина образуется один из главных гормонов роста растений — ауксин (Р-индолилуксусная кислота).
Рассмотренные связи дыхания и других процессов метаболизма растения не являются постоянными, раз навсегда данными. Они возникают и нарушаются под влиянием как внутренних особенностей растения, так и внешних условий. При неблагоприятных условиях эти нарушения могут быть значительными и даже летальными.
9.Количественные показатели газообмена
Дыхательный контроль. Возрастание функциональной активности клеток сопровождается усилением дыхания. В значительной степени это достигается благодаря механизму дыхательного контроля, или акцепторного контроля дыхания. Дыхательным .контролем называют зависимость скорости потребления 02 митохондриями от концентрации ADP, который служит акцептором фосфата при окислительном фосфорилировании. В условиях полного сопряжения транспорта электронов по ЭТЦ с синтезом АТР интенсивность дыхательного процесса в митохондриях зависит от концентрации ADP или, точнее, от отношения действующих масс АТР-системы: [ATP]/[ADP] [PJ. Причем неорганический фосфат обычно присутствует в достаточном количестве и не является ограничивающим фактором. В клетке, находящейся в состоянии покоя, это отношение достаточно велико, так как почти весь ADP фосфорилирован. При увеличении функциональной активности клеток АТР расходуется на энергозависимые процессы, в результате чего возрастает концентрация ADP, а это в свою очередь приводит к повышению скорости переноса электронов и интенсивности окислительного фосфорилирования. Важно подчеркнуть, что в данном случае уровень ADP регулирует интенсивность транспорта электронов и окислительное фосфорилирование не как аллостерический фактор, а как субстрат фосфорилирования.
Более полно состояние адениннуклеотидной системы выражается отношением, получившим название энергетического заряда:
[АТР] + у2 [ADP] [АТР] + [ADP] + [AMP] '
который характеризует меру заполнения всей адениннуклеотидной системы высокоэнергетическими фосфатными группами.
10.Регуляция процесса дыхания. Зависимость дыхания от внутренних факторов
Эффект Пастера. Уровень 02 в тканях влияет не только на интенсивность дыхания, но определяет и величину расходования дыхательных субстратов, на что впервые обратил внимание J1. Пастер. В его опытах с дрожжами в присутствии 02 снижались распад глюкозы и интенсивность брожения (уменьшалось количество спирта и выделяемого С02), но одновременно наблюдался интенсивный рост биомассы дрожжей вследствие усиления использования Сахаров на синтетические процессы. Торможение распада Сахаров и более эффективное их использование в присутствии кислорода получило название «эффекта Пастера».
Механизм эффекта Пастера состоит в том, что в присутствии 02 интенсивно идущий процесс окислительного фосфорилирования конкурентно уменьшает количество молекул ADP, вступающих в гликолиз (на нужды субстратного фосфорилирования). По этой причине, а также из-за тормозящего действия АТР (синтез которого резко возрастает в аэробных условиях) на фосфофруктокиназу, скорость процессов гликолиза в присутствии 02 снижается. Избыток АТР может способствовать и ресинтезу глюкозы из части молекул пирувата, образующегося в ходе гликолиза. Без кислорода не функционируют цикл Кребса и ПФП и, следовательно, клетки не получают многих промежуточных соединений, необходимых для синтеза клеточных структур. В присутствии 02 все эти циклы работают. Увеличение концентрации молекул АТР в условиях аэробиоза также способствует синтетическим процессам.
11.Зависимость процесса дыхания от факторов внешней среды
Концентрация 02. Процесс дыхания связан с непрерывным потреблением кислорода клетками и тканями растений. В то же время окислительные превращения субстратов при дыхании включают не только аэробные, но и анаэробные процессы (гликолиз, брожение). Поэтому интересен тот факт, что при снижении парциального давления 02 с 21 до 9% интенсивность дыхания тканей растений меняется незначительно. При 5% 02 у молодых растений начинает несколько снижаться поглощение 02 и в меньшей степени изменяется выделение С02. Как отмечалось, одна из важнейших конечных оксидаз аэробного дыхания — цитохромоксидаза — обладает высоким сродством к кислороду и может функционировать при его низких парциальных давлениях.
Эти факты можно объяснить как тем, что в эволюции дыхательные системы растений формировались в низкокислородных условиях, так и тем, что возможности функционирования аэробной дыхательной цепи при низких парциальных давлениях 02 — один из важных адаптивных признаков, элемент надежности энергетики растения. Это тем более важно, что газовый состав внутренних тканей органов растений может отличаться от атмосферного. Так, в листовой паренхиме сахарной свеклы содержание 02 меняется в течение суток от 7,1 до 17,4%, а С02 - от 0,9 до 5,1%; в мякоти зрелых яблок содержится 7,5 С02 и 13,9% 02, а плодов лимона — 8,5 С02 и 11,5% 02.
У растений, обитающих в условиях систематического затопления (болотные и др.), существует ряд приспособлений к перенесению недостатка кислорода: развитие аэренхимы, способность использовать кислород нитратов («нитратное» дыхание), различные способы устранения избытка продуктов брожения (этанола, молочной кислоты), а также механизмы их использования на нужды обмена веществ.
Избыток кислорода в тканях растений может возникать лишь локально. В атмосфере чистого кислорода дыхание растений снижается, а при длительном его действии растение погибает. Эффект избытка 02 связан с усилением в клетках свободнорадикальных реакций, повреждением мембран вследствие окисления их липидов и в конечном итоге — с нарушением многих процессов обмена веществ.
Диоксид углерода. Повышение концентрации С02 как конечного продукта дыхания приводит к снижению интенсивности дыхания. При повышении концентрации С02 тормозятся реакции декарбоксилирования и активность сукцинатдегидрогеназы и, следовательно, уменьшаются выделение С02 и дыхательный коэффициент. При этом наблюдается закисление тканей — ацидоз, что может приводить к вредным последствиям. С02 хорошо растворяется в липидах и таким образом может воздействовать на мембраны. По-видимому, он обладает некоторым наркотическим действием. Предполагается, что С02 способен регулировать метаболизм растений в анаэробных условиях. У листьев ингибирующее действие высоких концентраций С02 на дыхание может быть связано с закрыванием устьиц в этих условиях. Повышенное содержание С02 в тканях семян, покрытых плотной оболочкой, — один из способов поддержания состояния покоя.
Температура. Дыхание, подобно другим ферментативным процессам, зависит от температуры. В определенных температурных границах эта зависимость подчиняется правилу Вант-Гоффа (скорость химических реакций удваивается при повышении температуры на 10 °С). В интервале температур от 0 до 20 °С <210 дыхания равен 2 — 3. При температурах выше 20 °С величина Ql0 может понижаться. Одна из причин этого — уменьшение растворимости 02 в жидкостях при повышении темпепатупы. ЭАсЬект темпепатупы проявляется обычно вместе с другими факторами. Например, в условиях высокой температуры сильнее подавляется дыхание при пониженном содержании 02 или повышенной концентрации С02 и т. д.
Для дыхания каждого вида растений и его органов существуют определенные минимальные (когда дыхание еще возможно), оптимальные и максимальные температуры. В отличие от фотосинтеза дыхание у зимующих растений наблюдается при очень низких температурах (у хвои ели и сосны при — 25 °С). Температурный оптимум его у большинства видов умеренных широт лежит в пределах 35 — 40 °С, т. е. на 5 — 10 °С выше, чем для фотосинтеза. Максимальные температуры (45 — 55 °С) определяются способностью белков к денатурации.
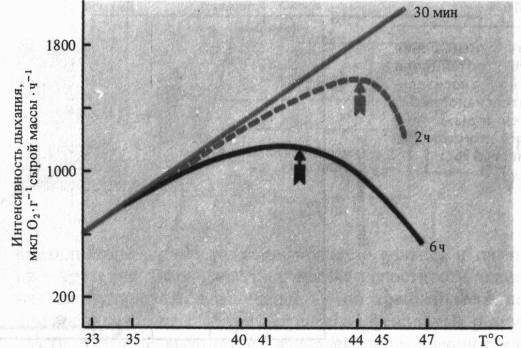
При рассмотрении влияния на дыхание крайних температур важно учитывать продолжительность действия этого неблагоприятного фактора. Так, при кратковременных экспозициях (30 мин) температурный оптимум дыхания листьев гречихи сахалинской может быть очень высок (> 47 °С), тогда как при двухчасовом действии оптимум смещается к 44 °С, а при шестичасовом—до 42 °С.
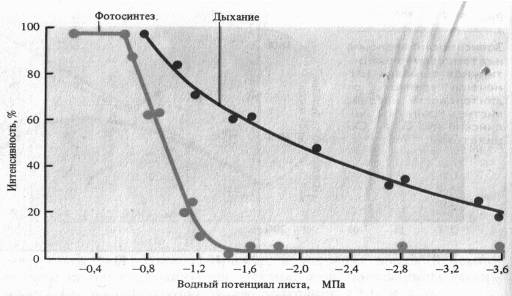
Водный режим. Изменение оводненности тканей растения отражается на интенсивности дыхания. В листьях проростков при быстрой потере воды вначале отмечается усиление дыхания — обычная кратковременная реакция на раздражение. При постепенном снижений оводненности этого не происходит. Продолжающийся водный дефицит вызывает постепенное снижение дыхания, причем более медленное, чем падение интенсивности фотосинтеза. Более раннее прекращение фотосинтетической активности при водном дефиците является следствием закрытия устьиц (уменьшение поступления С02 в лист) и потери тургорного давления в клетках.
Необходимость определенной степени оводненности тканей для процесса дыхания отчетливо проявляется при изучении дыхания семян. Дыхание воздушно-сухих семян (10—11% Н20) ничтожно мало. При повышении влажности семян до 14—15% дыхание возрастает в 4 —5 раз, а при дальнейшем повышении оводненности семян до 30 — 35% дыхание увеличивается в тысячи раз. При этом важную роль играет температура, при которой хранятся семена: при 0— 10°С влияние влажности на интенсивность дыхания значительно меньше, чем при 18 — 25 °С. Резкий подъем дыхания набухших семян сопровождается значительным выделением тепла, что может привести к их перегреву («самовозгоранию») при хранении.
Реакция растительных тканей на потерю воды зависит также от возраста, физиологического состояния, экологической принадлежности организма.
Минеральные вещества. Добавление раствора солей в воду, на которой выращивают проростки, обычно усиливает дыхание корней. Этот эффект получил название «солевого дыхания». Полагают, что солевое дыхание отражает усиление обмена веществ, необходимое для энергетической поддержки активного транспорта ионов корнем. Однако до конца это явление не изучено. Отмечены случаи, когда увеличенная интенсивность дыхания сохранялась после удаления солей. В тканях других органов этот эффект удается получить не всегда.
Свет. Действие света на дыхание зеленых органов растений из-за методических трудностей изучено недостаточно, так как одновременно с дыханием в них осуществляется фотосинтез. Освещенность, при которой интенсивность фотосинтеза равна интенсивности дыхания (по уровню поглощенного и выделенного С От), называется компенсационным пунктом.
При работе с незелеными растениями или с незелеными органами растений возможно непосредственное изучение действия света на дыхание. Дыхание (поглощение 02 и выделение С02) незеленых тканей активируется светом коротковолновой части спектра — ближним ультрафиолетом (380 нм), синими и зелеными лучами (400 — 500 нм) .
В области 380 — 600 нм лежат максимумы поглощения каротиноидов, флавинов и всех цитохромов дыхательной цепи митохондрий. Усиление дыхания может быть связано с прямым действием света на эти компоненты окислительно-восстановительных систем клеток.
Остается нерешенным вопрос о характере взаимодействия процессов фотосинтеза и дыхания в зеленых клетках в ходе фотосинтеза. Обсуждаются две точки зрения об использовании АТР, образующегося при фотофосфорилировании. Согласно одной из них на свету хлоропласта большую часть синтезируемой при фотофосфорилировании АТР используют на синтез углеводов, отдавая на нужды клетки лишь небольшую ее часть. АТР в цитоплазму поставляют митохондрии, которые используют ассимиляты. В темноте хлоропласта сами нуждаются в поступлении энергии из цитоплазмы.
По другой гипотезе АТР фотосинтеза на свету обеспечивает энергией не только фиксацию С02, но и другие энергозависимые процессы в клетке. Окислительное фосфорилирование в митохондриях на свету резко замедляется, а цикл Кребса ускоряется, так как его интермедиаты необходимы для функционирования хлоропластов.
Список использованной литературы
1.В.В. Полевой Физиология растений М.: Высшая школа 1989г.
2.Н.И. Якушкина Физиология растений М.: Просвещение 1993г.
Похожие работы
... ; обнаружены все ферменты, катализирующие превращение этих кислот; показано, что малонат — ингибитор сункцинатдегидрогеназы — тормозит окисление пирувата и резко снижает поглощение 02 в процессах дыхания у растений. Большинство ферментов цикла Кребса локализовано в мАТФиксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрии. Энергетический выход цикла Кребса, его ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... цитохромоксидаза, подтверждением чего служит также максимальная величина подавления дыхания азидом . Соотношения указанного типа не являются, однако, общей закономерностью, поскольку в ряде случаев максимальная интенсивность дыхания обнаруживается в клетках зоны растяжения. Такая закономерность выявляется при пересчете на белок, а также на единицу кислотонерастворимого фосфора. Вполне вероятно, ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев