Навигация
Вторым по значению методом, которым пользуются в
27392
знака
0
таблиц
0
изображений
2. Вторым по значению методом, которым пользуются в
настоящее время для дифференциации отдельных видов
гемоглобина, является хроматография 0.
Особенно хорошие результаты получается при употреблении в
качестве адсорбирующего вещества ионообменной смолы амберлита
и ионообменный декстрановый гель. На рис. 17 даны
характеристики различных типов гемоглобина, полученных при
применении амберлита при рН=6,0.
3. Для разграничения некоторых видов гемоглобина
пользуются также их растворимостью в некоторых растворителях 0.
Наиболее известным тестом этой группы является проба
Итано для доказательства наличия HbS. При этой пробе нам
служит то обстоятельство, что редуцированный HbS осаждается в
2,24 m буфере, в противоположность другим типам гемоглобина
(рис. 18). Проба эта имеет значение в особенности для
дифференцирования HbS и HbD, потому что, как видно из рис. 18,
HbS и HbD обладают одинаковой электрофоретической и
хроматографической подвижностью.
4. Для отличия HbA от HbF пользуются, как было
подчеркнуто выше, устойчивостью при денатурации растворами
натриевой щелочи. Это известный в истории метод, которым
Кербер в 1886 году дифференцировал HbA и HbF.
5. Гемоглобины группы F (HbF, Hb Феллас, Hb Александра и
Hb Бартс) отличается от других гемоглобиновых типов и по своей
характерной триптофановой полосе при 289,8 нм
ультрафиолетового спектра. Гемоглобины, обладающие группой М,
не имеют абсорбционной полосы при длине волны 630 нм, но зато
показывают увеличенную абсорбцию при 600 нм.
6. " Отпечатковый метод 0" .Дело касается важнейшего метода
установления "первичной структуры" гемоглобина при различных
гемоглобиновых типах. Исследуемый гемоглобин гидролизуют
трипсином, при чем полипептидные цепи глобиновой молекулы
распадаются на большое число пептидов. Пептидную смесь
подвергают электрохроматографии на бумаге, т.е. в одном
направлении проводится электрофоретическое, в другом
хроматографическое разделение. Получаются характерные для
отдельных типов гемоглобинов электрохроматограммы, по которым
их можно точно различить (рис. 19). Определение
аминокислотного состава отдельных пептидов дает возможность
первичную структуру глобина соответствующего гемоглобинового
типа. Делая аналогию с соответствующей по сложности и точности
криминалистической техникой для изучения отпечатков пальцев
рук, он был назван "пальцеотпечатковым" ("fingerprint")
методом.
7. Для определения состава полипептидных цепей в
каком-нибудь гемоглобиновом типе можно воспользоваться и так
называемым " 2рекомбинационным 0" или " 2гибридизационным 0" методом.
Если смешать известный и неизвестный гемоглобин при рН 4,3,
они диссоциируют полумолекулами, состоящими из соответствующих
пар полипептидных цепей. После нейтрализации раствора
полипептидные пары снова комбинируются в целые гемоглобиновые
молекулы, при чем могут получится и новые "гибридные"
гемоглобиновые молекулы. Их идентифицирование
электрофоретическим способом или хроматографией позволит
сделать заключение о полипептидной структуре неизвестного
гемоглобинового типа. Этот метод предназначен также
преимущественно для научных исследовательских целей.
8. 2 Иммунологические методы.
9. Кроме вышеуказанных методов при дифференциации
отдельных типов гемоглобина пользуются также 2различиями в
2кристаллическом строении, изоэлектрической точке и т.д.
10. Разработаны также методы 2цитологического определения
типа гемоглобина в эритроцитах на мазке крови. Так наличие HbF
в эритроцитах можно доказать путем обработки кровяного мазка
лимоннокислой буферной смесью с рН 3,2-3,6. При этих условиях
HbA извлекается и эритроциты, в которых он преобладал,
остаются только в виде эритроцитных теней, тогда как HbF
сохраняется и эритроциты, содержащие преимущественно этот тип
гемоглобина, сохраняют свое содержание.[8]
_ГЕМОГЛОБИН ПРИ СЕРПОВИДНОКЛЕТОЧНОЙ АНЕМИИ
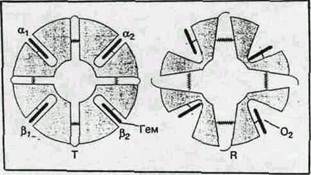
В гемоглобине S остаток Glu А2(6)бета замещен на Val.
Остаток А2 (Glu или Val) располагается на поверхности молекулы
гемоглобина и контактирует в водой, и замещение полярного
остатка Glu на неполярный Val приводит к появлению на
поверхности бета-субъеденицы "липкого участка". Этот липкий
участок присутствует как в оксигенированном, так и в
дезоксигенированном гемоглобине S (в гемоглобине А
отсутствует). На поверхности дезоксигенированного гемоглобина
существует комплементарный участок, способный прочно
связываться с липким участком бета-субъединицы, тогда как в
оксигенированном гемоглобине этот участок маскируется другими
группами (рис. 20). Когда гемоглобин S переходит в
дезоксигенированное состояние, его липкий участок связывается
с комплементарным участком на другой молекуле
дезоксигенированного гемоглобина. Происходит полимеризация
дезоксигемоглобина S и его осаждение в виде длинных волокон.
Волокна дезоксигемоглобина S механически деформируют
эритроцит, предавая ему серповидную форму, что приводит к
лизису клеток и множеству вторичных клинических проявлений.
Таким образом, если бы можно было можно поддерживать
гемоглобин S в оксигенированном состоянии или по крайней мере
свести к минимуму концентрацию дезоксигенированного
гемоглобина S, то нам удалось бы предотвратить полимеризацию
дезоксигенированного гемоглобина S и образование "серповидных"
клеток. Ясно, что полимеризации подвержена Т-форма гемоглобина
S. Интересно отметить (хотя в практическом плане это
малосущественно), что ферри-ион метгемоглобина А остается в
плоскости порфиринового кольца и тем самым стабилизирует
R-форму гемоглобина. То же относится и к гемоглобину при
серповидноклеточной анемии: гемоглобин S в ферри-состоянии
(метгемоглобин S) не подвержен полимеризации, поскольку он
стабилизирован в R-форме.
В дезоксигемоглобине А тоже имеется рецепторный участок,
способный взаимодействовать с липким участком
оксигенированного или дезоксигенированного гемоглобина S
(рис.20), но присоединения "липкого" гемоглобина S к к
дезоксигемоглобину А недостаточно для образования полимера,
поскольку сам дезоксигемоглобин А липкого участка не содержит
и не может связывать следующую молекулу гемоглобина.
Следовательно, связывание дезоксигемоглобина А с R- или
Т-формой гемоглобина S перекрывает полимеризацию.
В результате полимеризации дезоксигемоглобина S
образуются спиральные фибрилярные структуры. При этом каждая
молекула гемоглобина контактирует с четырьмя соседними
молекулами (рис. 21). Образование подобных трубчатых волокон
ответственно за механические нарушения в содержащем их
эритроците: он приобретает серповидную форму (рис. 22),
становится подверженным лизису в момент прохождения им щелей в
синусоидах селезенки.
_ТАЛАССЕМИИ
Другая важная группа нарушений, связанных с аномалиями
гемоглобина - талассемии. Для них характерна пониженная
скорость синтеза альфа-цепей гемоглобина (альфа-талассемия)
или бета-цепей (бета-талассемия). Это приводит к анемии,
которая может принимать очень тяжелую форму. В последние годы
достигнут ощутимый прогресс в выяснении молекулярных
механизмов, ответственных за развитие талассемии.[9]
СПИСОК ЛИТЕРАТУРЫ
1. Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл, Биохимия
человека, том 1, "Мир", Москва 1993г., стр.52
2. И.Тодоров, Клинические лабораторные исследования в
педиатрии, "Медицина и физкультура",София 1968г., стр. 278-281
3. Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл, Биохимия
человека, том 1, "Мир", Москва 1993г., стр.56-59
4. И.Тодоров, Клинические лабораторные исследования в
педиатрии, "Медицина и физкультура",София 1968г., стр.283-284
5. тоже стр.293
6. тоже стр.285-286
7. тоже стр.293-304
8. Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл, Биохимия
человека, том 1, "Мир", Москва 1993 г.6 стр. 60-62
_II. Дополнительная литература
Dean J., Schechter A.N. Sickle-cell anemia: Molekular
and lubar basis of therapeutic approaches. (3 parts),
N.E.Med., 1978, 299, 752, 804, 863.
Klotz I.M., Haney D.N., King L.C. Ritional approaches
chemotherapy: Antisickling agents, Sience, 1981, 219
Похожие работы
... находящегося в плацентарной крови. Однако после рождения ребенка HbF утрачивает свою функцию; обладая более высоким сродством к кислороду, он высвобождает меньшее его количество в тканях. У гемоглобина есть еще одна немаловажная функция: он ускоряет транспорт углекислого газа от тканей к легким. Гемоглобин связывает углекислый газ сразу после высвобождения кислорода; примерно 15% углекислого газа ...
... особенностей взаимного варьирования содержания белковых фракций в анализируемых образцах нами был проведен многомерный количественного содержания белков эритроцитарных мембран человека. Была построена матрица фенотипических корреляций количественного содержания гемоглобина и основных белков мембран эритроцитов. Данная матрица представлена в таблице 2. Из нее видно, что полученные коэффициенты ...
... метод предназначен также преимущественно для научных исследовательских целей. 8. 2 Иммунологические методы. 9. Кроме вышеуказанных методов при дифференциации отдельных типов гемоглобина пользуются также 2различиями в 2кристаллическом строении, изоэлектрической точке и т.д. 10. Разработаны также методы 2цитологического определения типа гемоглобина в эритроцитах на мазке крови. Так ...
... активности клеток в кроветворных органах и тканях. Приобретенная аутоагрессивная гемолитическая анемия: в селезенке происходит разрушение и растворение (лизис) эритроцитов. Биохимия и патобиохимия печени. Рассмотрим основные представления о биохимических процессах, протекающих в печени. На пути между кишечником и внутренней средой организма – системой крови и лимфы – находится печень. В ...
0 комментариев