Род
Campanula L. Систематическая
характеристика
L. speculum-veneris Dur
Общие
проблемы нодальной
анатомии
Проблемы
нодальной
анатомии семейства
Campanulaceae
Формирование
лакуны начинается
до образования
контактов между стеблем
и пазушной
почкой. Лакуна
формируется
с двух сторон
Методика
преподавания
темы “Внутреннее
строение стебля
в курсе ботаники
средней школы”
Результаты
методического
исследования
Навигация
Общие проблемы нодальной анатомии
Нодальная анатомия некоторых представителей семейства Campanulaceae Juss
148749
знаков
3
таблицы
4
изображения
1. Общие проблемы нодальной анатомии.
Нодальная анатомия - наука, изучающая картины сосудистой связи листьев и стебля, которые зависят от филлотаксиса, величины и строения листьев, типа жилкования (Эверт, Айкуорн, Рейвн, 1990, по Анели,1962). Она зародилась во Франции в XIX веке (Негели, 1858; Де-Берн, 1880; Грави, 1891, по Анели,1962). Большой вклад внес Д. Синнот (1914). Он первый начал данные нодальной анатомии в классификации растений. Среди зарубежных ученых можно выделить также А. Д. Имса (1964) и К. Эзау (1969). В отечественной науке ведущее место занимают школы ученых В. Г. Александрова и О. Н. Фадкевич (Александров и Александрова, 1920, 1932, 1954; Фадкевич, 1947; Анели, 1959, 1962; Завалишина, 1947, 1966; Василевская, 1947; Кондратьева-Мельвиль, 1956; Тахтаджян, 1962, 1964).
Строение проводящей системы оси побега покрытосеменных сложно и имеет определенный “рисунок”, который повторяется от узла к узлу (Имс, 1964). На поперечном срезе строения узла еще более усложняется благодаря отходящим тяжам проводящей системы - листовым следам, которые отходят в лист.
Термин “листовой след” трактуется по разному. Одни ученые принимают определение, предложенное Линье (Lignier, 1888 - цит. по Завалишиной, 1986), согласно которому под листовым следом понимается вся совокупность отходящих к листу пучков. Другие называют листовым следом каждый из проводящих пучков листа, входящих в центральный циллиндр, ориентируя это тем, что число таких пучков на лист представляет собой существенный признак (Имс, 1964).
Рисунок 1. Листовой след. (Лотова, 2000 стр. 161):
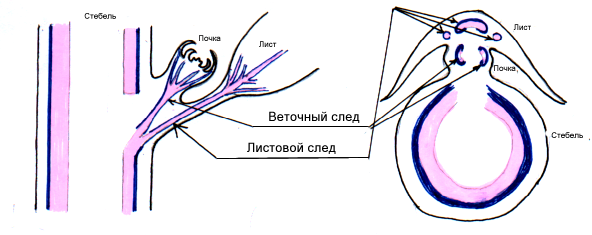
Один лист может иметь один или более листовых следов. Границы листового следа не могут быть строго определены, т. к. он простирается от стели к листу. В коре след может разветвляться или соединяться с другими следами (Имс, 1961). В проксимальной части (по отношению к центральному циллиндру) на различных уровнях следы объединяются с циллиндром стели или стеблевыми пучками в соответствии со свойственными растению числом следов и филлотаксисом. Как показывает подсчет междоузлий, пересекаемых следом, перед вхождением в стелу стебля, листовые следы бывают разной длины - менее одного междоузлия у хвойных, от одного до тридцати пяти междоузлий у различных двудольных (Эзау, 1980). Продольные пучки, от которых отходят листовые следы, относятся к стеблевым пучкам, называемым иначе каулиновыми (caulis - стебель) пучками (Эзау, 1980). Вследствие общности происхождения, никаких фундаментальных различий между листовыми следами и стеблевыми пучками нет, соответствующие термины имеют описательное, топографическое значение.
Но структура листовых следов и стеблевых пучков в деталях неодинакова. На поперечном срезе различается, что листовые следы, как правило, круглые и имеют большое количество ксилемы, чем стеблевые пучки. Как видно на продольном срезе, листовой след расширяется выше места ответвления от стеблевого пучка. Другой характерной чертой листовых следов у представителей многих таксонов является присутствие в сосудистых тканях паренхимных клеток, которые имеют выросты клеточной оболочки и служат передаточными клетками (Эзау, 1969).
В области узла, где листовой след ответвляется от проводящего цилиндра стебля, в последнем формируется зона паренхимы, называемая листовой щелью (лакуной или прорывом). Листовые щели могут быть различной протяженности как в латеральном, так и в вертикальном направлении. В верхних частях стебля травянистых растений листовые щели шире, чем в нижних (Александров, 1966). Если центральный цилиндр имеет много вытянутых вдоль оси межпучковых зон, щели сливаются с ними и говорить о границах лакуны можно только условно. В дополнение к листовым следам в зоне узла могут быть видны проводящие пучки, которые связывают главный стебель с веткой. Эти тяжи называют следами веток. В действительности, веточный след является листовым следом, а именно листовым следом двух (у однодольных одного) первых веточных листов, называемых профиллами. (Эзау,1969). Почку с главной осью стебля связывают два, по одному к каждому профиллу - листовых следа, состоящих из одного или большего числа пучков. Ветка и несущий лист имеют одну общую щель (Эзау, 1969; Имс, 1961). Число листовых следов и листовых щелей различно у разных растений, а так же может варьировать вдоль стебля одного и того же растения (Имс, 1961).
По Эзау (1969) если от узла отходит более одного листа, узел характеризуется только по одному из них. Анели (1962) для характеристики мутовчатых узлов вводит термин “кратерность”, под которым подразумевается совокупность всех каумефолиаров (листовых следов) одного листа, вступающих через общий канал - кратер - в центральном циллиндре. Число кратеров равно числу листьев в мутовке. Кратерность является одной из важнейших характеристик мутовчатого узла.
Для понимания строения узла большое значение имеет понимание процесса закладки и дифференциации тканей в онтогенезе растения. Вопрос о характере развития проводящей системы был поставлен еще в первой половине XIX века. Одни ученые были сторонниками акропетального развития проводящей системы, то есть от корня в стебель, а оттуда в листья (Бекетов, 1840; Де-Бари, 1885). Другие являлись сторонниками базипетального развития проводящей системы под влиянием листьев, т. е. развития в нисходящем направлении (Годишо, 1841; Ганштейн, 1858; Грави, 1935, по Александрову,1932). Некоторые исследователи считали, что проводящая система развивается одновременно в обоих направлениях. К настоящему времени все больше и больше укрепляется мнение, что проводящая система развивалась в нисходящем направлении под влиянием листовых и веточных следов (Радкевич, 1947; Камилова и др., 1961; Василевская, 1940; Александров 1932, 1954, 1966; Эзау, 1969). Годишо в 1841 годы первым открыл нисходящее развитие проводящих элементов от листа вниз. Это открытие позднее легло в основу теории листовых следов, которая гласит, что развитие проводящей системы стимулируется листьями. Основные положения теории листовых следов следующие:
1. Проводящие пучки возникают в зоне контакта листа и стебля, т. е. в узлах. Побег с самого начала развивается на основе взаимодействия оси, листьев и пазушных почек.
2. Дифференциация проводящих элементов совершается по мере вытягивания междоузлия в нисходящем направлении, пучки уменьшаются в объеме книзу.
3. Заложенная у основания листьев и стимулируемая их ростом (путем гормонального влияния) Проводящая система представляет собой совокупность листовых следов.
Более детально развитие проводящей системы растения описывается так (по Александрову, 1966).
При дифференциации тканей коры и сердцевины, в основании листьев остается тяж меристемы, называемый продесмогеном. Он продолжается в стебель, и там имеет форму кольца в результате слияния дуг продесмогена всех зачатков листьев (остаточная меристема по Эзау, 1969). Выше уровня закладки листьев никакой дифференциации сосудистых тканей не обнаруживается. В связи с дальнейшим ростом зачатка листа паренхимные клетки продесмогена делятся и вытягиваются, превращаясь в тяж паренхимных меристоматических клеток - прокамбий. Основание прокамбия начинается с более старой зоны листа - его подножия и распространяется вверх по зачатку листа и вниз по стеблю. Клетки прокамбия в той же последовательности превращаются в проводящие ткани. Если отрезать какой-либо начинающий развиваться лист (листовой зачаток) и спустя некоторое время после операции сделать поперечный разрез стебля ниже места прикрепления ранее срезанного зачатка листа, можно убедиться, что сосудисто-волокнистые пучки, связанные с жилками отрезанного молодого листа, имеют ненормальный вид или совсем не развиваются. Степень дегенерации проводящего пучка стебля зависит от того, связан ли пучок с одним отрезанным развивающимся листом или при посредстве анастомозов соединен с пучками, идущими от других листьев, а также от того, на какой стадии развития был отрезан начинающий развиваться лист. Итак, в своем развитии сосудисто-волокнистые пучки стебля связаны с развитием определенных листьев. Поэтому сосудисто-волокнистые пучки стебля называют листовыми следами. До тех пор, пока сосудисто-волокнистый пучок, следуя вниз по стеблю, не сливается с другими пучками, его можно назвать индивидуальным листовым следом.
Когда же он соприкасается с другими листовыми следами и сливается с ними, образуется синтетический листовой след. Таким образом, Александров расширяет понятие “листовой след”, включая под это определение пучки, лежащие вдоль оси стебля (Эзау разделяет понятие “листовой лед” и “стеблевой пучок”). Это мнение можно считать наиболее категоричным выводом теории листовых следов.
Под воздействием развивающегося листа участки прокамбия, расположенные в стебле ниже места прикрепления листа, постепенно превращаются в элементы сосудисто-волокнистого пучка. Новые сосудисто-волокнистые пучки, дифференцирующиеся из межпучкового камбия в сформировавшихся уже участкам стебля, представляют собой нижние концы листовых следов, возникших при образовании новых листьев, зачатки которых появились на конусе нарастания стебля при дальнейшем развитии побега. Сосудистые пучки как бы проникают из пучка в стебель. Дифференцируясь дальше звено за звеном вниз по стеблю, вновь образовавшиеся листовые следы встречают на своем пути более старые, идущие от ниже расположенных листьев, и сливаются с ними. Таким путем создается целостная сеть сосудистых пучков стебля и листа. Аналогично описывает данные процессы Эзау (1969).
По Александрову (1936) образование лакуны также происходит благодаря влиянию листовых зачатков (“Об образовании щели в центральном цилиндре стебля двудольных растений. Бот.журн. 1936, т.21, № 5, стр. 525): “Вся совокупность структурных явлений, происходящих при образовании листовой щели, убеждает в том, что камбий по существу представляет собой меристематическую ткань без определенной направленности к образованию определенных тканей. Лишь под влиянием определенных раздражений, идущих от листа, камбий образует соответствующие элементы центрального цилиндра (Александров, 1922). По-видимому, даже само направление распространения исходящего раздражения имеет немалое значение для дифференциации того или иного анатомического элемента (Александров и Абесадзе, 1934). Возможно, что когда взаимно стакиваются раздражения, исходящие от двух листовых органов, расположенных в различных ярусах, как это имеет место в районе каждого узла (диссонирующие раздражения), из камбия образуются паренхимные элементы, которые и создают ткань листовой щели. Раздражение же, идущее более обособленно и при этом в одном направлении (консонирующие раздражения) дифференцирует из камбия нормальные анатомические элементы центрального цилиндра”.
Нодальная анатомия находит все более широкое применение в разных областях ботаники. В первую очередь стоит упомянуть о значении нодальной анатомии для филогении и систематики. Анели (1962) писал, что имеющимся в то время филогенетическим схемам для многих семейств довольно трудно установить родственные отношения и историю, т. к. исследователи лишены возможности опираться в своей работе на единую детально разработанную филогенетическую систему. Возникает необходимость применения дополнительного метода, который базировался бы на консервативных признаках и который позволил бы внести ряд корректив, необходимых для объяснения многих неясных положений в систематике и филогении покрытосеменных. На основе комплексных морфолого-анатомических данных вопросы эволюции покрытосеменных, несомненно будут разрешаться более объективно. Проводящая система растений является одним из наглядных и убедительных наследственных признаков, содержащих в достаточной степени элементы консервативностей и прогресса. Особое внимание заслуживает первичная проводящая система побега, в частности, проводящая система, связывающая лист и стебель, точнее кауфолиарная система (от слова “кауфолиар” - листовой след) черешка и узла. Важно, что текстура кауфолиарной системы остается, в принципе, стабильной в разных экологических условиях.
Синнот (1914) изучил строение узла у различных видов и пришел к выводу, что типы анатомии узла очень константны и обычно характерны для целых семейств или даже порядков покрытосеменных. Синнот начал активно использовать данные нодальной анатомии в установлении естественной классификации растений. Примечательно, что его данные в последствии, за редким исключением, подтвердились работами Анели (1962).
Анели (1962) выделяет два типа семейств по строению кауфолиарной системы: монотекстурные (для них характерен один тип строения узла) и политекстурные (имеют две-три типовые текстуры кауфолиарной системы). Эволюция развития родов в политекстурных семействах пошла по различным направлениям и их можно считать эволюционно продвинутыми. Кроме того, по строению узла выделяются семейства однолакунные, трехлакунные, многолакунные и поликауфолиарные (соответствуют политекстурным). Так же семейства отличаются по количеству пучков в лакунах. Эти данные кроме систематики, могут использоваться в диагностике растений (Анели, 1962; Эзау, 1962).
Для построения филогенетических связей и направлений эволюции большое значение имеет установление примитивных типов узлов. По этому поводу было написано много работ и выдвигались разнообразные версии. Как правило, эволюция узла рассматривалась в связи с эволюцией листа. Синнот и Бейли (1914) указывали на примитивность трехлакунного узла (часто связанным с простым листом, имеющим пальчатое жилкование, который рассматривался как примитивный). Обоснование примитивности трехлакунного узла находили в том, что такое строение узла особенно характерно для многих примитивных форм (у Archichlamybeae с преобразованием в многолакунные типы у Epacriolaceae, Platonaceae, Araliaceae, Umbeliferae, Polygonaceae, Meliaceae и в редуцированные типы у Centrospermae, Myrtiflorae и большинства Metachlamybeae. У некоторых Archichlamybeae наблюдается редукция числа следов в пределах семейства, у некоторых - увеличение числа следов). Кроме того, узлы с тремя следами обнаружены у проростков двудольных, взрослые растения которых имеют узлы со многими следами. Однодольные обычно являются многолакунными, однако и их проростки часто имеют узлы с тремя следами, а их плодолистики, обычно обладающие многими следами, могут иметь у редуцированных форм три следа (Имс, 1961). Эта точка зрения на примитивность трехлакунного типа узла получила вскоре почти полное произнание, но уже к концу 40-х годов и особенно в 50-х годах она начинает пересматриваться.
На основании изучения анатомии у Magnoliaceae и других апокарпных двудольных Озенда (1949, по Александрову,1966) пришел к выводу, что примитивным типом узла является не трехлакунный, а многолакунный. По его мнению, три типа нодальной структуры покрытосеменных - регрессивный ряд, который начинается с многолакунного типа и кончается однолакунным.
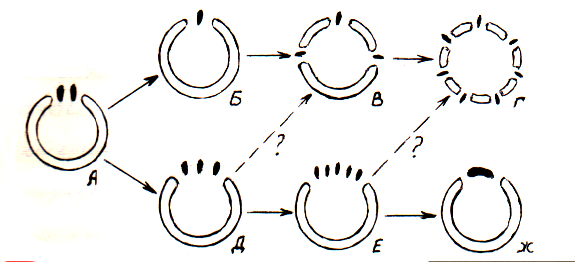
Наконец, в 50-х годах ряд авторов (Марсден, Бейли, 1955; Конрайт, 1955 и др., по Александрову,1966) выступили за примитивность однолакунного узла с двумя дискретными листовыми следами, названного четвертым типом узла. Новая концепция эволюции нодальной структуры покрытосеменных, нашедшая поддержку со стороны других анатомов (Гиффорд, 1959 по Александрову,1966; Эзау, 1960; Имс, 1964). Такой листовой след характерен для многих голосеменных и папоротников. Он так же встречается у некоторых двудольных (Lactoris, Austobaileya, Monimiaceae и др.). У семядолей и профиллов так же часто встречаются парные или слившиеся медианные следы. Парные следы, по-видимому, более обычны в узлах семядолей, чем в узлах вегетативных листьев. Двойные медианные пучки часто обнаруживаются в плодолистиках представителей семейств, далеко отстоящих друг от друга в системе (Ericaceae, Coryophyllaceae). У некоторых свободных плодолистиков рыльца двулопастные и проводящие пучки раздваиваются. Двойные пучки были описаны у многих таксонов при изложении теории полиморфизма плодолистиков. Двухследовые тычинки необычны, но обнаружены как у представителей продвинутых таксонов, так и у примитивных. Типы узлов с нечетным числом следов, столь характерным для покрытосеменных вообще, по Имсу (1961) возникли путем слияния двух следов примитивного узла. Слияние двух следов однолакунного узла приводит к образованию узла с одним следом; добавление пар боковых следов, образующих свои листовые щели, формирует 3-х и многолакунный узел с нечетным числом следов (рис. 2 В). Продвинутые типы однолакунных узлов с единственным следом развились путем утраты боковых следов и путем латерального слияния трех и более следов и их прорывов (рис. 2 Д, Е, Ж).
Рисунок 2. Схема эволюции узла по Имсу (Имс,1961 стр.318):
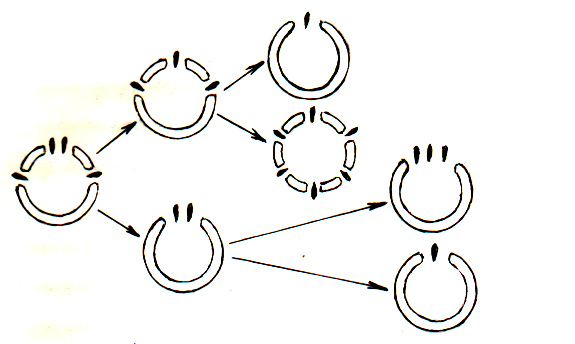
Тахтаджян выдвинул свою гипотезу на развитие узловой структуры. В своей работе “Основы эволюционной морфологии покрытосеменных”, стр. 192, он пишет: “Несомненно правы Марсден, Бейли, Имс и другие, когда они утверждают, что эволюция шла от четного числа листовых следов к нечетному: с другой же стороны, число лакун всегда нечетное (1, 3, 5, 9 и т. д.). Из этого следует, что исходным типом узла мог быть только узел с нечетным числом лакун и четным числом листовых следов. Это мог быть или однолакунный узел с двумя листовыми следами (4-ый тип); или же узел с тремя или несколькими (5, 7 и больше) лакунами, от медианной лакуны которого отходило 2 листовых следа (назовем его 5 типом).
Данные по анатомии узла наиболее примитивных групп покрытосеменных убеждают меня в том, что это скорей 5-й тип, чем 4-ый. В пользу этого предположения говорит тот факт, что почти все семейства Magnolia sensu Stricto обладают 3-х или многолакунным узлом. Черезвычайно важно так же, как показали наблюдения Свали (1949), средняя лакуна в узле Magnoliaceae и Degeneria имеет двойной листовой след. По своей нодальной анатомии с Magnolia ales имеет много общего порядок Trochodendrales. Узлы с 3, 5 или реже 7 и больше лакун характерны также для таких относительно примитивных порядков как Nymphaeales, Puperales, rosales, dilleniales. Важно также отметить, что двойной листовой след наблюдается не только у растений с одной лакуной. Как листовой след однолакунного узла, так и листовой след медианной лакуны трех-многолакунного узла имеет иногда более или менее ясно выраженную двойственную природу, что уже давно было установлено для семядолей (Томас, 1967), а в ряде случаев и для средней жилки листа взрослого растения (Имс, 1931). Особенно интересна Degeneria, в трехлакунном семядольном узле которой ясно видны два дискретных листовых следа, отходящих от медианной лакуны. Все эти данные приводят к выводу, что первичным типом узла у покрытосеменных является узел с тремя или большим числом лакун. Определить более точно исходный тип узла в настоящее время не представляется возможным, но если мы примем во внимание, что трехлакунный тип полностью отсутствует у голосеменных, а многолакунный имеется у Cycdaceae и Cynetum, мы должны сделать вывод, что проводящая структура примитивных покрытосеменных была скорее многолакунного, чем трехлакунного типа.”
Рисунок 3. Схема эволюции узла по Тахтаджяну
(Тахтаджян, 1964 стр.198):
По Анели (1962) развитие первичных покрытосеменных пошло по крайней мере, в начале по 4-м направлениям. Первое - платановые - Platonites (многолакунность со множеством пучков в лакуне), второе - магнолиевые - Magnolites (многолакунность со множеством пучков в средней лакуне и по одному пучку в краевых лакунах), третье - лавровые - Laurites (однолакунность со множеством пучков) и четвертое - ореховое - Juglandites (трехлакунность со множеством пучков в каждой лакуне). Исходными примитивными растениями нужно считать трехлакунные, многолакунные и однолакунные. Эволюция пошла по линии многопучковости в каждой лакуне и сокращению количества пучков в лакуне. Вполне возможно, из многолакунных и трехлакунных растений эволюция шла так же по линии сокращения количества лакун (Анели, 1962)
С. Ф. Завалишина (1966) считает, что “нельзя определять примитивность или прогрессивность в строении узла, исходя только из числа лакун и числа листовых следов. Многолакунность и многопучковость точно также как однопучковость и однолакунность при детальном изучении формирования могут относиться к совершенно различным типам узла”.
Ряд анатомов придает филогенетическое значение, кроме числа лакун и следов, ряду других признаков. Болеепримитивной считают проводящую систему, когда пучки листового следа долго сохраняют свою индивидуальность. Чем больше угол, под которым входят пучки листового следа, тем примитивнее узел (Александров, 1954; Завалишина, 1966).
Данные нодальной анатомии можно использовать в смежных отраслях ботаники для разрешения некоторых практических и теоретичеких вопросов. Пока намечены следующие отрасли: гибридизация, прикладная систематика, вегетативное размножение растений путем окоренения стеблевых черешков, идентификация и диагностика лекарственного сырья (Анели, 1962).
Нодальная анатомия быстро развивается. К 60-м годам (Анели, 1952, Анели, Штрамберг, 1964 по Александрову,1966) узловая анатомия содержит по крайней мере 6 кардинальных признаков:
1. Тип внедрения или приложения листовых следов в первичной коре узла.
2. Количество лакун.
3. Многопучковость или однопучковость.
4. Тип индивидуального следования листовых следов.
5. Скорость внедрения листовых следов в центральный цилиндр.
6. Степень меристематизации и витальности узловой сферы.
К ним прибавляются новые (Анели, 1962); кратерность, широта потенциальной части узла, широта реактивной части узла, широта контактной части узла, общее количество проводящих пучков во всех листьях одного кратера, общее количество проводящих пучков во всех кратерах одного узла, протяженность акцентированных листовых следов в последующих узлах и междоузлиях, типовая картина размещения листовых следов в отношении центрального цилиндра стебля и др. Совершенствуются методы нодальной анатомии. Проводящую систему обычно изучают, зарисовывая положение проводящих пучков на серийных срезах и затем реконструируя по этим рисункам объемное изображение. Этот метод анализа сложен даже по отношению к узлам, имеющим всего один или несколько следов в лакуне, к однодольным он вообще не применим. Существенный прогресс в понимании организации проводящей системы был достигнут при использовании микросъемки серийных поперечных срезов стебля. Кинокадры проецируются на чертежную бумагу так, чтобы можно было измерить смещение пучков на последовательных срезах. Метод дает возможность быстро проанализировать трехмерную структуру стебля.
0 комментариев