Навигация
О передаче метаболических сигналов в системе нейрон – нейроглия
33355
знаков
1
таблица
1
изображение
2. О передаче метаболических сигналов в системе нейрон – нейроглия.
(Н. Г. Алексидзе Тбилисский гос. Университет, Тбилиси, СССР)
Гипотеза А. И. Ройтбака об участии глиальных клеток в замыкании временных межнейрональных контактов стимулировала исследования по биохимии нейронно – нейроглиальных взаимоотношений В настоящее время всеми признается что нейрон-нейроглия является функциональной единой системой, однако материальная сущность передатчиков сигнала от нейрона на клетки глии и многие вопросы, связанные с его реализацией в метаболические процессы, остаются открытыми.
Было высказано предположение, что биохимический цикл глиального обеспечения функции нейронов осуществляется путем обратной метаболической связи при непосредственном участии в качестве передатчиков сигнала нейромедиаторов, К+, аммиака и др. соединений. Предпосылки для такого заключения имелись как в нейрохимической, так и в физиологической литературе, но требовалось коррелятивное сопоставление физико-химического состоя мембран глии с биохимическими процессами в них в условиях моделирования возбуждающего и тормозного состояния нейрон – нейроглиального комплекса.
Исходя из вышесказанного мы предприняли исследование участия К+, нейромедиаторов и аммиака в передаче метаболических сигналов от нейрона на клетки нейроглии в условиях их целостного состояния, на уровне изолированных единичных нейронов и глиального скопления или же обогащенных нейронами и глиальными клетками фракций.
МЕТОДИКА.
Объектом исследования служили беспородные белые крысы и кролики. Нейроны и глиальные клетки выделяли из срезов головного мозга кроликов методом Хидена, обогащенные нейронами и глиальными клетками фракции — из коры больших полушарий крыс и кроликов по прописи Роуза в модификации. Скорость поглощения кислорода изолированными нейронами в глиальными клетками измеряли методом поплавка в модификации Хидена и Пигона, в опытах с обогащенными фракциями был использован полярографический метод определения дыхания.
Результаты.
Влияние К+ на скорость потребления клетками глии и нейронов.
Впервые, об особой чувствительности глиальных клеток к К+ указал Куффлер с сотрудниками. Позже этот вывод был обоснован биохимически датским ученым Хертцом. Однако анализ его результатов был затруднен, так как изолированные нейроны были получены в основном из коры головного мозга кошек, а скопления глии - из коры мозга крыс.
Кроме того, имелись определенные недостатки и в методике исследования. После разработки метода дубль поплавка удалось показать стимулирующее влияние К+ на дыхание клеток глии вестибулярного ядра Дейтерса, а нейроны, даже в специальных опытах с предварительно измененным содержанием К+ в инкубационной среде от 5мМ до 60мМ, не обнаруживали существенных различий в скорости поглощения кислорода.

Рисунок 1. Влияние К+ на дыхание нейрона(2) и нейроглии(3) латерального вестибулярного ядра Дейтерса кролика. Стрелками указан момент добавления К+ в инкубационную среду. 1-контроль, без нервных клеток.
В опытах с обогащенными нейронами и глиальными клетками фракциями при избытке К+ Бредфорд и Роуз наблюдали примерно равное усиление их дыхания, а согласно данным Хультборна и Хидена скорость поглощения кислорода изолированными нейрональными клетками возрастала примерно в 2 раза. Вотличии от результатов Бретфорда и Роуза, работая с фракциями обогащенными нейронами и глиальными клетками, Халиаме и Хамбергер подтвердили результаты Хертца и наши об особой чувствительности клеток глии к К+. В связи с такими разногласиями о действии К+ на нервные клетки, мы провели анализ экспериментальных условий описанных выше опытов. Как выяснилось, при получении обогащенных нейронами и глиальными клетками фракций в градиенте фикола и сахарозы в среде Роуза концентрация, К+ была 100 мМ, а в среде Хамбергера, Хертца и нашей, концентрация К+ не превышала 5 мМ. Из данных литературы известно, что повышение концентрации К+ до 100 мМ и выше вызывает моментальное и обычно необратимое изменение цитоплазмы глиальных клеток и ее сморщивание. Высокие концентрации К+ в среде культивирования нервных клеток вызывали увеличение объема глиальных клеток и уменьшение содержания в них сухого остатка. В нейронах такие изменения не были найдены. Следовательно, можно было предположить, что низкая чувствительность глиальных клеток к К+ в опытах Брэдфорда и Роуза в условиях повышенного содержания К+ в среде их выделения обусловлена предварительной деполяризацией и сенсибилизацией мембран глии. Это было доказано экспериментально.
При замене К+ ионами натрия в среде выделения нервных клеток резко возрастает чувствительность обогащенных глиальными клетками фракций к К+ усиление дыхания, по отношению к контролю (1,5 мМ К+), составило примерно 90%. Таким образом, была выяснена причина разногласия относительно чувствительности нервных клеток к К+ и высказано предположение об участии К+ в передаче метаболического сигнала от нейрона на клетки нейроглии.
Ацетилхолин как передатчик метаболического сигнала в системе нейрон-нейроглия.
При изучении возможной роли ацетилхолина (АХ) в качестве передатчика метаболического сигнала в нейрон-нейроглиальной системе мы исходили из следующих фактов 1) ацетилхолин освобождается при возбуждении и вызывает сдвиги в мембранной активности глии;
Таблица 1
Влияние ацитилхолина(АХ) на скорость поглощения кислорода (ОАХ) обогащенными клетками глии фракций при разных соотношениях К+/АХ. О0-скорость поглощения кислорода без АХ. Концетрация АХ-10-5г/мл
| К+ мМ | Скорость поглощения кислорода мкА О2/мин | ||||
| Оо | % | Оах | % | В % к Оо | |
| 5 мМ | 6,82±0,45 | 100 | 8,06±0,81 | 100 | 118,2 |
| 40 мМ | 10,21±0,62 | 161,1 | 12,87±0,42 | 159,7 | 126,1 |
| 60 мМ | 13,45±0,73 | 197,2 | 12,5±0,54 | 155,1 | 92,2 |
2) под влиянием АХ изменяется активность ряда ферментов обмена углеводов, липидов, белков, нуклеиновых кислот и т.п.
В связи с вышеизложенным, мы предприняли исследование наличия связи между изменением мембранной активности клеток глин и углеводным обменом в них при воздействии АХ. При этом особое внимание обращалось на соотношение К+ к АХ (К+—5 мМ/АХ — 10-5 г/мл; К+- 40 мМ/АХ—10-5 г/мл; К+—60 мМ/АХ—10-5 г/мл).
Было установлено (табл. I), что при концентрации К+ 5 мМ скорость поглощения кислорода клетками глии в присутствии АХ возрастает на 18%. При более высокой концентрации К+ (40 мМ) эффект АХ усиливается и достигает 26%, а при концентрации К+ 60 мМ стимулирующий эффект АХ на дыхание элиминируется, а по сравнению с контролем даже проявляется тенденция к торможению. Стимулирующий эффект АХ на скорость поглощения кислорода полностью исчезает в присутствии аптихолинэргического агента — атропина. Этот факт указывал на существование холинэргического рецептора на мембранах глии, что в настоящее время является хорошо доказанным экспериментально. Подтверждается существование холинэргического механизма регуляции дыхания глиальных клеток, где роль информатора сигнала может выполнить возбуждающий нейропередатчик АХ.
ГАМК как передатчик метаболического сигнала в нейрон-нейроглиальной системе.
Из данных литературы известно, что под влиянием ГАМК изменяется мембранная активность глии и стимулируются окислительные процессы в нервной ткани. Следовательно, можно было допустить, чти и ГАМК может претендовать на роль метаболического сигнала. С целью решения данного вопроса в качестве объекта были взяты нервные клетки ядра Дейтерса кролика, где функцию нейропередатчика выполняет ГАМК. Изменения в содержании ГАМК вызывали введением ГАМК и фармакологических веществ (гидроксиламин, тиосемикарбазид), действие которых связано с обменом ГАМК. Об изменении метаболической активности изолированных нейронов и клеток глии судили по сукцинатоксидазной (СО) активности (СОА), которая является удобным тестом для оценки функционального состояния нервных клеток.
Было установлено, что в зависимости от уровня содержания ГАМК в головном мозгу активность СО меняется реципрокно: ГАМК подавляет активность фермента в нейронах, а в глии напротив—стимулирует. Под влиянием гидроксиламина по сравнению с нормой более чем в 2 раза возрастает САО нейронов, в нейроглии—подавляется. Тиосемикарбазид также стимулировал СОД в нейронах, но не оказывал влияния на активность фермента в клетках глии. Учитывая разнонаправленность действия ГАМК, гидроксиламина и тиосемикарбазида па количественное распределение ГАМК в головном мозгу и результаты влияния ГАМК на окислительное фосфолирование было сделано заключение, что в регуляторных механизмах окислительных процессов нервных клеток значение имеет не общее содержание ГАМК в мозгу, а ее распределение во внутри- и в внеклеточном пространстве. Следовательно, и ГАМК может выполнить функцию передатчика сигнала в нейрон-нейроглиальной системе.
Аммиак как передатчик метаболического сигнала в нейрон-нейроглиальной системе.
Уровень аммиака в головном мозгу является одним из показателей функционального состояния ЦНС. Как было установлено, обмен аммиака находит свое отражение в мембранной активности клеток глии, что послужило основанием изучения его возможной роли в передаче информации о функциональном состоянии нейронов на перинейрональные клетки. С целью биохимического обоснования такого механизма мы исследовали дыхание обогащенных клетками глии фракций в опытах in vitro в зависимости от концентрации аммиака в среде инкубации. В качестве субстрата дыхания использовали глутамат и глутамин. Дыхание клеток глии в присутствии глутамата служило контролем. В опытных вариантах образование глутамата, субстрата дыхания, происходило в результате распада глутамина. Следовательно, при такой постановке опытов, критическими были: величина активности глутаминазы глии и скорость освобождения аммиака глутамата.
![]()
![]()
![]()
Рис. 2 Скорость поглащения кислорода глиальными клетками в присутствии глутаминовой кислоты (1) и глутамина (2).
Предварительные опыты по изучению глутаминазной активности глии показали, что она является аллостерическим ферментом высокой степенью кооперативности, следовательно требовалось графическое сопоставление скорости образования аммиака в инкубационной среде и скорости поглощения кислорода глиальными клетками. Как видно из рисунка 2, спустя одну минуту после добавления глутамина в инкубационную среду, в период максимального усиления дыхания, количество аммиака составляет 0,64 мкМ, через 4 мин, когда проявляется тенденция угнетения дыхания— 1.40 мкМ а на 9-й мин, при торможении дыхания на 60%—3,20 мкМ. В опытах с глутаматом (контроль) нам не удалось обнаружить достоверных изменений в продукции аммиака и, следовательно, дыхание глиальных клеток во времени возрастало линейно.
Суммируя вышеизложенное, мы считаем, что аналогично К+ АХ и ГАМК, аммиак также может участвовать в передаче метаболического сигнала от нейрона на нейроглиальные клетки.
Механизм инактивации нейропередатчиков глиальными клетками.
Исходя из того факта, что нейропередатчики могут выступать в роли переносчиков метаболических сигналов в нейрон-нейроглиальной системе, возникает вопрос о необходимости их инактивации глиальными клетками. В настоящее время установлено, что глиальные клетки обладают способностью инактивировать нейропередатчики на уровне плазматической мембраны и внутриклеточно. Примером первого пути является гидролиз АХ глией без предварительного его захвата. Клетки глии характеризуются высокой ацетил- и бутирилхолинэстеразной активностью и легко могут устранить излишки АХ. Продукт гидролиза АХ холин, который обладает слабым холинэргическим эффектом, устраняется клетками глии механизмом захвата высокого сродства. На примере ГАМК было показано, что в клетках глии имеется две системы его захвата: с высоким (Км = -31 ± 7 мкМ) и низким (Км--123±10 мкМ) сродством. Выявлены также механизмы активного захвата дофамина (Км -0,07± 0,001 мкМ) и серотонина (Км —0,083±0,002 мкМ). Дальнейшая судьба инактивации серотонина в клетках глии заслуживает особого внимания в связи с его отрицательным влиянием на синтез белков. Нам удалось установить, один из возможных механизмов инактивации серотонина в клетках глии путем синтеза глюкуронида серотонина, последний в отличие от серотонина отличается более чем в 1000 раз меньшей биологической активностью.
Таким образом, выясняется, что относительно всех кандидатов, Претендующих на информативную роль в передаче метаболических сигналов, в клетках глин имеются мощные механизмы устранения их хеморецептивного воздействия на мембрану.
Похожие работы
... общей пользой, но могло служить причиной смут, если монархом, например, Иваном IV Грозным, овладевали своеволие и личные страсти. Носителем «русскости» и добродетели в труде Глинки выступало простонародье, избегнувшее иностранного влияния и не руководствующееся в своих поступках эгоистическими интересами. Глинка избегал исторической критики источников, отдавая предпочтение прямому мифотворчеству, ...
... о традиции пасторалей и эклог (Давид в юности — пастух), о сентиментально-романтической образности. Мотив «желания славы», особенно остро воспринимающийся поколением, победившим Наполеона, ощутим в следующей строке — «О мне узнают ли народы?». Особая роль в создании образа младенчества — детства принадлежит стихотворению «Умащение ног» (1840-е) разворачивающему евангельский эпизод. Автор так ...
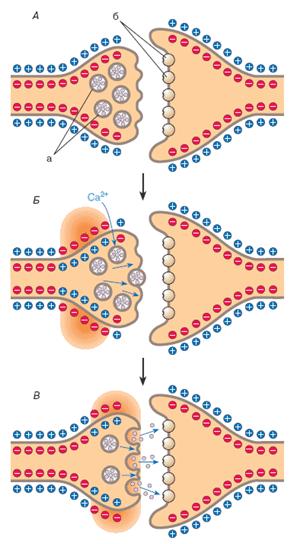
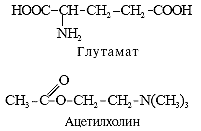
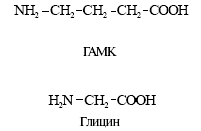
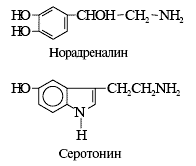
... ). Таблица 1. Структура низкомолекулярных нейтротрансмиттеров Характер действия Основная функция возбуждение ингибирование Нейромедиаторы Нейромодуляторы Нейромедиаторы Главные медиаторы головного мозга — аминокислоты. К возбуждающим относятся глутамат и аспартат. При освобождении в синапс (см. рис. 1, В) они через ионотропные рецепторы ( ...
... . Обзор работ по этим ферментам, который будет представлен ниже, является свидетельством большого интереса к пептидгидролазам нервной ткани нелизосомальной локализации, и вместе с тем это лишь первые шаги в выяснении функциональной роли этой группы пептидгидролаз. Характеристика протеолитических ферментов нервной ткани нелизосомальной локализации и их биологическая роль Пептидгидролазы ...
0 комментариев