Навигация
Возможная роль глиальных клеток в обеспечении нейронов АТФ. (Л. М. Чайлахян Институт проблем передачи информации АН СССР, Москва, СССР)
33355
знаков
1
таблица
1
изображение
3. Возможная роль глиальных клеток в обеспечении нейронов АТФ. (Л. М. Чайлахян Институт проблем передачи информации АН СССР, Москва, СССР)
В общей проблеме о функциональной роли нейроглии существует важный и интересный вопрос—являются ли глиальные клетки источником энергии для нейронов? Он возникает в связи с тем, что глиальные клетки, с одной стороны, не уступают нейронам по интенсивности энергетического обмена, в частности, но окислительному фосфорилированию, т. е. в производстве АТФ, но, с другой стороны, должны потреблять гораздо меньше энергии, чем нейроны, так как электрически пассивны. Для обоснования подобной точки зрения важно достаточно аккуратно сравнить энергетические потребности для поддержания ионных градиентов у нейронов и глиальных клеток. В настоящем сообщении сделаны такие количественные оценки, результаты которых позволяют сформулировать гипотезу о возможной роли глиальных клеток в обеспечении нейронов АТФ.
В первую очередь нужно оценить необходимые энергетические затраты нейрона для поддержания ионных градиентов в покое и сравнить их с таковыми у глиальных клеток. Для последующих расчетов на основании литературных данных была приняты следующие геометрические и электрофизиологические параметры для обобщенного нейрона и глиальной клетки.
Геометрические параметры нейрона: объем нейрона принимался равным объему шара диаметром 30н м - что соответствовало величине Он=1.4.10-8 см3, а площадь поверхности (Sн)-соответствовала увеличенной в 5 раз поверхности такого шара, что составляло Sн=1.4.10-4 см3.
Геометрические параметры глиальной клетки: объем главной клетки (Оr) принимался равным объему шара с диаметром 14нм, что соответствовало величине Оr=0,14.10-8см2, а площадь поверхности (Sr) соответствовала увеличенной в 5 раз поверхности такого шара, что составляло Sr=0.3.10-4 см2.
Электрофизиологические параметры нейрона и глиальной клетки: мембранный потенциал у нейрона в покое-Vмн= -70мв, у глиальной клетки Vмг= -89мв, потенциалы равновесия по ионам калия (Vк) и ионам натрия (VNA), а также удельные проводимости поверхностной мембраны у нейрона и глиальной клетки не отличались и принимались-Vk=-90мв, VNA=-60мв, gm=10-3 с/см2/так как проводимость поверхностной мембраны в основном определяется ионами калия, то принималось-gm=gk. Кроме того принималось, что у глиальной клетки отсутствует электрогенная Na, К-помпа. Решающие доводы в пользу последнего допущения были представлены на симпозиуме «Функции нейроглии» в Тбилиси в докладе Р. Г. Гроссмана. Было показано, что инъекция ионов натрия в глиальные клетки не приводит к появлению какой-либо заметной гиперполяризации, что свидетельствовало бы об электрогенной помпе, как это было показано в сходных опытах на нейронах моллюска.
Исходные предпосылки для расчетов. На основании принятых электрофизиологических параметров, соответствующих большому количеству исследований, при использовании известного уравнения Гольдмана—Ходжкина—Катца легко показать, что отношение проницаемостей для ионов натрия (РNA) и калия (Рк) у нейронов примерно на 1,5 порядка выше, чем у глиальной клетки-у нейрона PNA/Pk=0,031, а у глиальной клетки РNA/Pk=0,001.
Для последующих расчетов использовали уравнения:
gk(Vk-Vми)=gNAн(VNA-Vмн) (2)
gk(Vk-Vмг)=gNAг(VNA-Vмг) (3)
которые отражают условия равновесия в состоянии покоя у рассматриваемых клеток, когда пассивный ток ионов калия наружу должен быть равен пассивному току ионов натрия внутрь. Уравнение для нейрона, строго говоря, выполняется, если Na, К-насос, как для глиальных клеток, электронейтрален, т. е. стехиометрический коэффициент для активных потоков ионов натрия наружу и ионов калия внутрь равен. Однако, поправка на электронность будет лишь увеличивать энергетические затраты нейрона на ионные потоки.
На основании уравнений [2] и [3] и принятых нами параметров для нейрона и глиальной клетки можно вычислить величины ионных токов для этих клеток на единицу поверхности клетки (см2) или веса (гр) и времени (секунды, часы, сутки). Знание электрохимических градиентов для ионов калия и ионов натрия позволяет от значений токов перейти к оценкам соответствующих энергий. Очевидно, что энергия, затрачиваемая на Nа, К-насосы, поддерживающая пассивные ионные потоки, должна быть не меньше оцениваемой нами описанным способом.
Результаты расчетов. Существенно оценить затраты энергии у разных тканей на единицу поверхности клеток, так как эти оценки непосредственно отражают интенсивности ионных потоков и не зависят от размеров и формы клеток.
Для нейрона в покое получено: 0,3*10-5вт/см2. Если принять, что частота работы нейрона в среднем составляет 15-30 импульсов в секунду, то сравнительные оценки по пассивным потокам у нервных клеток в покое я при возбуждении дают основания предполагать, что затраты на сохранения ионного гомеостаза при такой работе нейрона могут увеличиваться в два-три раза, т. е. достигать 1 • 10-5вт/см2.
Интересно заметить, что вычисленные нами затраты энергии для идеализированного нейрона на единицу поверхности удивительно хорошо совпали с экспериментальными данными Конноли и Крейнфельда, полученными для гигантского аксона кальмара на основании измерений потребления кислорода – 0,5*10-5 вт/см2.
Для глиальных клеток вычисленные энергетические затраты на ионный гомеостаз были такие: 0,15*10-6 вт/см2. Видно, что энергетические затраты на единицу площади на работу Na, К-насоса у глиальной клетки должны быть в 20—60 раз меньше, чем у нейрона.
Знание отношения S/O у нейрона (104) и у глиальной клетки (2,14*104) позволяет перейти от затрат на работу насосов на единицу поверхности к затратам на единицу массы соответствующей ткани. Вот эти цифры: для нейрона в покое -0,3*10-1вт/гр, для работающего нейрона -l*10-1вт/гр, для глии -0,32*10-2вт/гр. Видно, что и в пересчете на единицу массы энергетические затраты у глиальных клеток на насосы в десятки раз меньше, чем у нейрона.
Сравнение энергетических затрат у нейрона на ионный гомеостаз и синтетические процессы. Представленные оценки могут не производить сильного впечатления, если думать, что затраты на ионный транспорт в нервной клетке составляют небольшой процент от всех суммарных энергетических затрат. Однако, видимо, это не так.
Для нервных клеток энергетические затраты на ионный гомеостаз составляют подавляющий процент во всем энергетическом балансе.
Для подтверждения этой точки зрения можно привести сравнительные оценки энергетических затрат у нейрона и глиальной клетки на ионный транспорт и синтетические процессы, в частности, на синтез белка. Примем, что интенсивность синтеза белка в нервной клетке такова, что за сутки происходит полное воспроизведение всех белков. Зная процентное содержание белка в нервных клетках (8% от веса клетки) можно оценить количество пептидных связей на единицу веса ткани. А знание необходимой энергии для синтеза одной пептидной связи (примерно гидролиз трех молекул АТФ до АДФ) позволяет оценить соответствующие энергетические расходы на воспроизведение белка за сутки.
Для нейрона в покое и при активации, а также для глиальной клетки выше уже приводились данные об энергетических затратах. Для удобства сравнения с затратами на синтетические процессы они также будут пересчитаны на сутки.
Вот эти цифры. На ионный транспорт энергетические затраты в сутки у нейрона в покое -2592вт/гр, при активности -5000 -7500вт/гр, у глиальной клетки -258 вг/гр, а на синтетические процессы у нейрона и глиальной клетки указанной выше интенсивности расходуется в сутки—61. гр.
Расчеты показывают, что на столь интенсивный синтез белка, как полное его воспроизведение за сутки, нервная клетка в покое должна затрачивать в 42 раза меньше энергии, чем на ионные насосы, а при активной работе в 123 раза меньше. Даже у глиальной клетки на ионные насосы затрачивается в 4,2 раза больше энергии, чем на столь интенсивные синтетические процессы.
Поистине, дорого стоит нервной ткани поддержание в боевой готовности натриево-калиевого механизма генерации и проведения нервного импульса — практически на это идут все энергетические затраты.
Все это означает, что если глиальные клетки в целом способны с такой же интенсивностью синтезировать АТФ как и нейроны, то АТФ у них должна быть в избытке. И из соображений целесообразности естественно предположить возможность прямого использования этого избытка нейронами.
Формулировка гипотезы. Можно предложить возможный путь потока АТФ из глиальных клеток в нейроны на основе уже известных механизмов. Этот путь должен состоять из двух этапов.
Первый этап — это выброс АТФ из глиальных клеток при их деполяризации ионами калия во время активации соседних нейронов (имеются убедительные данные, что при калиевой деполяризации глиальные клетки активно секретируют в межклетники ряд еще неидентифицированных соединений).
Второй этап — это поступление АТФ из межклетников в пресинаптические окончания по механизму пиноцитозного поглощения (в пресннатических окончаниях показано существование процесса обратной секреции—типа пиноцитоза).
С точки зрения этой гипотезы нейроглия является общим распределенным энергетическим резервуаром, снабжающим нейроны универсальным биологическим топливом — АТФ. Активность того или иного нейронного пула сразу же приводит к калиевой деполяризации глиальных клеток, окружающих эти нейроны. Они начинают секретировать АТФ в межклетники, а оттуда через активированные пресинаптические окончания эта АТФ может поступать по механизму пиноцитозного поглощения в нейроны. Таким образом, при реализации подобной возможности видна большая целесообразность во взаимодействии нейронов и глиальных клеток—поток АТФ из глиальных клеток в нейроны четко регулируется самой нейронной активностью: чем активнее работает нейрон, тем больше АТФ в него будет поступать.
Важно также заметить, что наличие щелевых контактов между глиальными клетками создает условия для эффективного диффузионного обмена АТФ глиальными клетками. Другими словами система глиальных клеток, окружающая нейроны, может в этой связи рассматриваться как единая непрерывная диффузионная среда, в которой могут осуществляться градиентные потоки АТФ в участки мозга с наибольшим потреблением АТФ, т. е. в места наибольшей нейронной активности. Таким образом, может происходить своеобразная кооперация глиальных клеток при обеспечении АТФ наиболее нуждающихся нейронов.
Высказанные соображения мало чего стоят, пока не будут получены прямые экспериментальные данные в пользу сформулированной гипотезы.
Заключение.
В заключение хочу обобщить все сказанное.
Во всех органах человеческого тела, кроме мозга, функционирующие клетки удерживаются вместе межклеточным веществом соединительной ткани. В нервной системе эту роль выполняет глия ( от греч. глия-клей), клетки которой образуются из общих с нейронами предшественниц на раннем этапе развития мозга. Глия создает опору для нейронов, объединяет отдельные элементы нервной системы, но, в то же время, изолируют друг от друга разные группы нейронов, а также большую часть их аксонов. Тем она формирует структуру мозга. Численность клеток глии превышает нейронов в мозгу приблизительно в 10 раз. Эти клетки отличаются друг от друга по внешнему виду и по выполняемой функции.
Самым распространенными среди клеток глии являются астроциты, например, в мозолистом теле они составляют 1/4 всех клеток глии. У астроцита неправельной, звездчатой формы тело с многочисленными и относительно длинными отростками, один из которых направлены к нейронам, а другие- к кровеносным капиллярам. Эти отростки расширяются на концах, образуя т. н. астроцитарную ножку. На поверхности капилляра отростки соседних астроцитов плотно смыкаются друг с другом и практически полностью обвертывают кровеносный сосуд. Подобная изоляция сосуда является одним из способов формирования гематонцефалического барьера- границы между кровью и нервной тканью, закрытой для многих находящихся в крови веществ.
Другие отростки астроцита почти целиком обертывают тела нейронов. Если нейрон возбуждается длительно, вокруг него повышается концентрация ионов калия, а это может уменьшить возбудимость соседних нейронов. Астроциты предупреждают такую возможность, поглощая излишки калия, тем самым они выполняют функцию буфера. Некоторые клетки глии при этом деполяризуются, а поскольку они связаны между собой щелевыми контактами, между деполяризованными и находящимися в покое клетками возникает ток. Это, однако, не приводит к возбуждению, так как в мембране клеток глии очень мало потенциалзависимых каналов для натрия и калия. Не смотря на, что повышение концентрации ионов калия у астроцитов изменяет некоторые их свойства, в настоящее время нет достаточных оснований считать их прямыми участниками переноса нервных импульсов.
Особую роль клетки глии выполняют, по-видимому, во время развития мозга. Некоторые их разновидности регулируют напровление перемищения нейронов в определенные регионы растущего мозга, а также напровление роста аксонов. Другие клетки глии возможно участвуют в питании нервных клеток путем регуляции кровотока, а тем самым транспорта глюкозы и кислорода.
Литература.
1.Костюк П. Г. «Структура и функция биологических мембран» М., «Наука» 1975 г.
2.Шульговский В. В. «Физиология ЦНС» Изд. Моск. универ. 1997 г.
3.Недоспасов В .О. «Физиология ЦНС» М.: ООО УМК «Психология» 2002 г.
Похожие работы
... общей пользой, но могло служить причиной смут, если монархом, например, Иваном IV Грозным, овладевали своеволие и личные страсти. Носителем «русскости» и добродетели в труде Глинки выступало простонародье, избегнувшее иностранного влияния и не руководствующееся в своих поступках эгоистическими интересами. Глинка избегал исторической критики источников, отдавая предпочтение прямому мифотворчеству, ...
... о традиции пасторалей и эклог (Давид в юности — пастух), о сентиментально-романтической образности. Мотив «желания славы», особенно остро воспринимающийся поколением, победившим Наполеона, ощутим в следующей строке — «О мне узнают ли народы?». Особая роль в создании образа младенчества — детства принадлежит стихотворению «Умащение ног» (1840-е) разворачивающему евангельский эпизод. Автор так ...
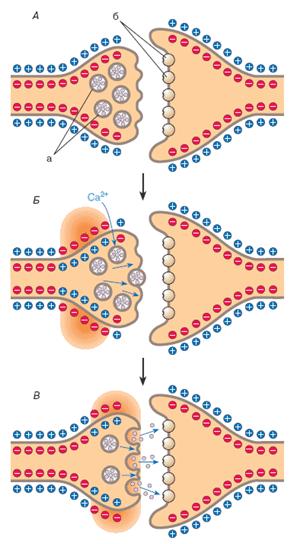
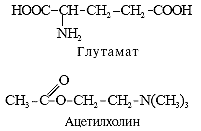
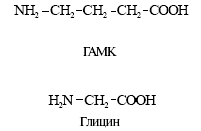
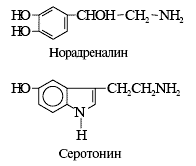
... ). Таблица 1. Структура низкомолекулярных нейтротрансмиттеров Характер действия Основная функция возбуждение ингибирование Нейромедиаторы Нейромодуляторы Нейромедиаторы Главные медиаторы головного мозга — аминокислоты. К возбуждающим относятся глутамат и аспартат. При освобождении в синапс (см. рис. 1, В) они через ионотропные рецепторы ( ...
... . Обзор работ по этим ферментам, который будет представлен ниже, является свидетельством большого интереса к пептидгидролазам нервной ткани нелизосомальной локализации, и вместе с тем это лишь первые шаги в выяснении функциональной роли этой группы пептидгидролаз. Характеристика протеолитических ферментов нервной ткани нелизосомальной локализации и их биологическая роль Пептидгидролазы ...
0 комментариев