Навигация
3. БИОСИММЕТРИЯ
2.3.1.БИОСИММЕТРИЯ СТРУКТУРНАЯ—МОЛЕКУЛЯРНАЯ
Содержание этого вида симметрии мы раскроем постепенно, переходя от нульмерных групп симметрии биомолекул к одно-, дву-, трехмерным. Из всех точечных групп симметрии для «мономерных» молекул наиболее характерны лишь две—п и п•т, при этом обычно п = 1, 2, ..., k, где k—величина небольшая. Поэтому наиболее распространенными группами здесь оказываются соответственно (1) и т, 2•т, 3•т... Первая характерна, например, почти для всех оптически активных — асимметрических — мономерных или олиго-сахаров, алкалоидов, многих аминокислот; вторые группы наиболее характерны для всякого рода оптически неактивных, часто запасных веществ. Однако недиссимметрическими группами иногда приходится описывать симметрию, подчас и чрезвычайно метаболически активных веществ (некоторые азотистые основания). Последнее обстоятельство резко ограничивает эмпирическое обобщение Г. Ф. Гаузе об обязательной диссимметричности метаболически активных соединений . Действительная картина здесь, таким образом, оказывается сложнее.
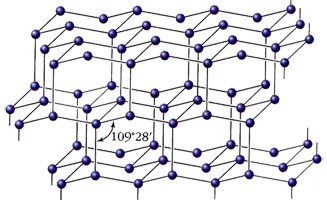
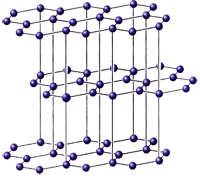
Аминокислоты, пуриновые и пиримиднновые азотистые основания, сахара и т.д., так или иначе химически взаимодействуя, «кристаллизуются» в полимерные, вытянутые в одном направлении цепные молекулы—белки, нуклеиновые кислоты, целлюлозу, крахмал гликоген и другие соединени. Выше мы видели, что цепные молекулы относятся к стержням, поэтому их симметрия должна исчерпываться всего 17 типами, охватывающими бесконечное множество видов симметрии. Однако учет характера взаимодействия между атомами «хребта» и боковых радикалов цепной органической молекулы, тенденций перехода в энергетически наиболее выгодное состояние и других факторов позволяет утверждать, что п природе наиболее часто должны встречаться ценные молекулы, принадлежащие к 13 группам симметрии стержней с N == 1 и к двум типам с винтовой осью «порядка» М — 8м н 5л»/2 .
Учет симметрии возможных конфигураций ковалентных связей главной оси— (2), (3), (3), (4) делает потенциально возможным для отдельных цепных молекул еще 30 групп, что дает всего 45 групп. Число «кристаллографических» групп цепных структур равно, как известно, 75. С возникновением живой природы число наиболее часто встречающихся групп резко уменьшается—до 4. Эти группы—диссимметрнческие: t,t/2, SM/2, где t—ось трансляции (обозначения международные). Например, целлюлоза и "полй-l-аланйн относятся к группе S2, полипептиды в конфигурации α-спирали — к S18/5.
Отдельные цепные молекулы могут давать образования из 2, З... цепочек. Если они связываются водородными связями, то их называют сложными, цепными молекулами; ван-дер-ваальсовыми (по принципу плотной упаковки; в первом случае он не выдерживается) —пучками; если сложная цепная молекула образована из химически различных единиц, то она называется комплексной цепной молекулой.
Сложные и комплексные цепные молекулы, пучки возникают главным образом в биосистемах; они оптически активны, представлены одной энантиоморфой. Поэтому они относятся к диссимметрическим группам стержней: tN, Sм N, tN/2, SMN/2. Однако учет меньшей устойчивости четверных и пятерных (чем двойных и тройных) цепей, спирализации как общего способа последовательной упаковки звеньев цепных молекул делает наиболее вероятным для сложных цепных молекул групп SM2, SM/2, SM3, пучков—Sм, Sм 3, комплексных цепных молекул—Sм2. Так, сложная цепная молекула ДНK относится к группе Sм/2, полиадениловая кислота — к S 2, полиинозиновая — к Sм 3, комплексная цепная молекула вируса табачной мозаики — к S49/3 . Последняя построена из уложенных по одноходовому пологому вунту белковых субъединиц, внутри которых идет цепочка РНК. На каждую субъединицу приходится три нуклеотнда; на три оборота молекулы приходится 49 белковых субъединиц. Другие примеры комплексной ценной молекулы—ДНК-протенды. Здесь полипептидная цепь белка обвивает молекулу ДНК по малой канавке. Так как эта цепочка одиночная, симметрия нуклеопротеи-да — Sм , хотя самой ДНК — Sм /2.
Другой способ объединения цепных молекул приводит к плоским двумерным фигурам — слоям. Причем сами цепные молекулы могут лежать в плоскости слоя или перпендикулярно ему (классический пример последних—парафины). Наиболее распространены слои первого рода, которые мы и рассмотрим.
Из 80 групп симметрии слоев для слоев из цепных молекул из-за особенностей их пространственного строения в первом приближении возможными оказываются 42 группы. Ограничения плотной упаковкидоводят их число до 19, а наиболее плотную упаковку фигур в слои позволяют всего 4 группы симметрии:
tt'с, tt'1, S2t, З2с. При переходе к биологическим, например мембранным, слоям число групп симметрии с 19 понижается из-за энантиоморфизма до 9: tt', tt'2, 2t, 21t, 2 (21) t, 222, 2122, 21212, 21(2)21 (2) 2 (NВ:S2=21). Классические примеры биологических слоев — складчатые слои полипептидных цепей, предложенные Паулингом и Кори3. Они могут быть параллельные и антипараллельные. Другой их пример — уже отмеченные мембранные слои.
При объединении полимеров в трех взаимно перпендикулярных направлениях пространства возникает ряд различных агрегатов, на одном конце которого идеальные кристаллы, на другом — совершенно аморфные вещества. Для живой природы характерны формы веществ, в той или иной мере отклоняющиеся от идеальных кристаллов и абсолютно аморфных тел.
Здесь, с одной стороны, наблюдается из-за богатства биополимеров Н-связями тенденция к самоагрс-гированию, п как следствие к образованию форм в той или иной мере упорядоченных—лент, складчатых кристаллов, кристаллов из слоев коротких цепных молекул и т.д. Так, хороню изученная кросс-β-конфигурация кератина является лентой из одной полипеп-тидной цепи, построенной по типу антипараллелыюго складчатого слоя. Другой пример. Как известно, в молекулах РНК в зависимости от ионной силы раствора и его температуры меняется число Н-связей, и это как следствие приводит к трем формам их существования: 1) нитям, 2) палочкам (аналогам лент), 3) клубкам.
С другой стороны, из-за больших и разнообразных длин цепных молекул, их гибкости, взаимодействия с соседями, спутывания, скручивания, образования прочных межцепных ковалентных связей между молекулами, например типа дисульфидных связей в каучу-ках, возникновение идеально упорядоченных во всем объеме кристаллов невозможно. Кроме того, такие квазикристаллы в свою очередь часто образуют в различной степени упорядоченные образования—мозаичные монокристаллы, текстуры, поликристаллы и т.д.
Особенности упорядочивания атомов и молекул в нуль-, одно-, дву-, трехмерные биологические образования дали повод Дж. Берналу выступить с идеей обобщенной кристаллографии, характерной прежде всего для живой природы. Она имеет дело уже не с «бесконечно» упорядоченными структурами, а со структурами с частичной упорядоченностью расположения атомов. Характернейшая ее особенность—учение о статистической — средней, наиболее часто встречающейся, вероятной — симметрии, с одной стороны, и нуль-, одно-, дву-, трехмерной «кристаллизации» (упорядоченности) — с другой .
Разумеется, такой переход к изучению кристаллов с нарушенной структурой стал возможным и исторически, и логически только после известного завершения учения об идеальных кристаллах. Он привел, как известно, к обоснованию молекулярной биологии. Опираясь на учение о последних и зная реальные кристаллы, стало возможным классифицировать различные типы нарушений. По Б. К. Вайпштсйну, основные их формы следующие: сдвиги, повороты, нарушения сетки и параллельности цепей; остальные их формы выводятся в результате комбинирования основных. К сказанному добавим, что в одних и тех же кристаллах во времени наблюдаются как процессы увеличения, так и уменьшения нарушений.
В заключение отметим резко проявляющееся в полимерных биомолекулах диалектическое единство асимметричного и симметричного, иррегулярного и регулярного строений. В белках естественного происхождения это проявляется, например, в асимметричности и нерегулярности их первичного строения (из-за уникальной линейной последовательности различных L и реже D аминокислот), в симметричности и регулярности их вторичного строения (часто из-за винтового закручивания всей или части полипептид-ной цепи), в резкой асимметричности и нерегулярности их третичного строения (из-за сложения полипеп-тидной цепи — поодиночке или в соединении с другими цепями в причудливые извитые трехмерные структуры, которые мы знаем как белковые молекулы), в столь же резкой симметричности и регулярности их четвертичного строения (из-за укладки идентичных белковых молекул в кристаллические и в квазнкри-сталлические структуры). Аналогично обстоит дело и с нуклеиновыми кислотами. В частности, первичная структура «молекулы жизни»—ДНК асимметрична и нерегулярна из-за уникальной последовательности нуклеотидов, в то время как ее вторичная структура явно симметрична и регулярна из-за винтовой закру-ченности двух ее цепей.
В итоге сравнения неживой и живой природы на молекулярном уровне неминуем эмпирический вывод о резкой диссимметризации, происшедшей при переходе от неживой природы к живой: 1) величина симметрии; 2) число возможных групп сильно уменьшаются; 3) наблюдается четко проявляющееся единство асимметрического и симметрического планов строения в основных «молекулах жизни», превращения типа «симметризация диссимметризация». Отсюда неизбежен вывод о специфическом характере биологической симметрии на молекулярном уровне.
Похожие работы


l Какие виды симметрий, встречаются в природе; l Как применяет красоту симметрии в своих творениях человек? Поэтому тему своего исследования я назвала «Симметрия — символ красоты, гармонии и совершенства». §2. Что такое симметрия? Ее виды в геометрии. О, симметрия! Гимн тебе пою! Тебя повсюду в мире узнаю. Ты в Эйфелевой башне, в малой мошке, Ты в елочке, что у лесной дорожки. С ...

... к старинным геохимическим циклам планеты). Осуществление завершающего звена, по В.И. Вернадскому, должно быть итогом переработки отходов и вышедшей из употребления продукции автотрофными, так как высшие формы живой материи - гетеротрофные - способны усваивать без ущерба для себя лишь химически чистые, однородные элементы. Хлорофильные растения и окисляющие бактерии должны поэтому выполнить роль " ...


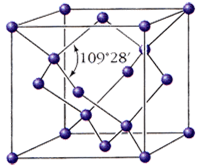
... требуется оценить отклонение от нулевого положения, например на руле грузовика или на штурвале корабля. Симметрия проявляется в многообразных структурах и явлениях неорганического мира и живой природы. В мир неживой природы очарование симметрии вносят кристаллы. Каждая снежинка- это маленький кристалл замерзшей воды. Форма снежинок может быть очень разнообразной, но все они обладают симметрией - ...
... по-видимому, прибавлялась только как искусственная роскошь к довольно узкому готовому миру вещей с их свойствами и силовыми взаимодействиями, их движениями и изменениями». Об определении категорий симметрии и асимметрии В настоящее время в науке преобладают определения указанных категорий на основе перечисления их важнейших признаков. Например, симметрия определяется как совокупность ...
0 комментариев