Навигация
× m®2 • т ® т®(1)®т®5 • т. ®(1)
90168
знаков
0
таблиц
3
изображения
8 × m®2 • т ® т®(1)®т®5 • т. ®(1).
К сожалению, филогенетическая эволюция симметрии по ходу отдельных ветвей древа жизни практически не изучена. Здесь явно необходимы планомерные исследования. Они могут привести к крупным открытиям и обобщениям.
Что касается характера изменения симметрии организмов в их онтогенезе, то помимо данных, приведенных выше, можно сослаться также и на интереснейшие результаты М.Д. Велибекова, полученные им при изучении ряда бобовых и других растений (подсолнечник, гречиха).
Он указывает, что обычно «в процессе развития состояние беспорядочной ориентации (чередования) и связи правых и левых метамеров, свойственное юве-нильному растению, заменяется константной; в дальнейшем (чаще на уровне цветок, плод, зародыш) ориентация и связь вновь становятся неопределенными» . Иначе, в их онтогенезе статистически начальная симметризация сменяется диссимметризацией, а последняя — снова симметризацией. Одновременно чаще одинаковые по частоте встречаемости и другим свойствам D и L фитоформы этих растений, их низкие полярность, целостность, пространственно-временная организованность, большие полиморфичность (информационная), энтропия, евклидовость в начале развития заменяются в ходе развития на неодинаковые по свойствам (в том числе встречаемости) D и L фитоформы, повышенные полярность, целостность, пространственно-временную организованность, пониженные полиморфичность, энтропию, евклидовость. В дальнейшем, в ходе отрицания отрицания, все эти показатели как бы снова возвращаются к исходным состояниям. И вот что замечательно: «Развитие информации, энтропии, пространственных характеристик циклично и обычно следует математическим закономерностям ряда золотого сечения».
Разумеется, отмеченные М. Д. Велибековым закономерности применимы не ко всем растениям, даже не ко всем бобовым. Возможно, в будущем будут точнее описаны и сами эти закономерности. Но при всем этом нельзя не отметить огромной ценности самого подхода, выдвинутых им характеристик, полученных данных, новизны развиваемого им направления в бносимметрике.
В итоге мы видим, что на морфологическом уровне: 1) величина симметрии организмов в ходе эволюции жизни закономерно в тенденции падает, образуя многоветвистые эволюционные ряды симметрии; и) на низших ступенях организмы представлены множеством видов симметрии. При этом их число много больше 32 — числа видов симметрии кристаллов. Однако к вершинам эволюционного древа число видов симметрии резко уменьшается, возникают многократно асимметризованные формы; 3) как и на уровне цепных молекул, появляются макробиоформы с запрещенными для кристаллов осями симметрии порядка 5, 7, 8, 9... Однако вопреки широко известному взгляду пятерная ось получает большое распространение не на всех, а лишь на определенных ступенях развития живого, как и двусторонняя симметрия т; 4) как в онто-, так и в филогенезе имеют место переходы типа диссимметризация « симметризация, причем процесс в целом сильно сдвинут в сторону диссимметриза-ции. Таким образом, и на макроуровне биологическая симметрия обнаруживает ряд специфических черт, что с новых сторон подтверждает положение В. И. Вернадского о специфическом характере биологического пространства.
Приведенные факты показывают, что воззрения на природу, построенные с позиций только одной из рассмотренных противоположностей, односторонни и в конечном счете неверны. Мир есть в рассматриваемом аспекте, насколько мы можем судить о нем с поправкой на сегодняшний уровень знаний, единство взаимоисключающих, обусловливающих, дополняющих, борющихся, переходящих друг в друга противоположностей, созидающих и одновременно нарушающих симметрию.
2.3.3. БИОСИММЕТРИЯ СТРУКТУРНАЯ — НЕКЛАССИЧЕСКАЯ
Приведенные в двух предыдущих параграфах данные позволяют сделать еще одно утверждение: на биообъектах реализована классическая симметрия абсолютно всех размерностей — точечная, линейная, плоская, пространственная. Однако не только классическая. Хотя биосимметрия с точки зрения различных неклассических теорий симметрии не изучена, ниже мы укажем по крайней мере на отдельные примеры реализации в живой природе главнейших из открытых в последние 50 лет симметрии.
Просто и l-кратно антисимметричны все те организмы и их части, которые обладают l+1=n дисс-факторами. Таковы, например, диссимметрические корни, стебли, листья, побеги, чашечки, венчики, цветки многих растений; внутреннее строение животных, множество оптически активных биологических соединений — Сахаров, аминокислот, белков, нуклеиновых кислот и т.д. Еще один конкретный пример, антисимметрии можно найти в работе Маизенхаймера, Нормана и Штербе. Они сообщают о существовании у некоторых рыб, например анаблепс, двух половых рас. Одна половая раса состоит из D самцов и L самок; другая, напротив, из L самцов и D самок. Оплодотворение оказалось возможным только в пределах своей I половой расы и невозможным между L самцами и L самками, а также D самцами и D самками.
С точки зрения учения о симметрии составляющие эти расы особи равны, симметричны (в известном приближении) друг другу в нескольких смыслах. Для более четкого их выявления примем только следующие обозначения: левое и правое по-прежнему будем обозначать буквами L и D, женский и мужской пол — знаками «+» и «—». Тогда мы придем к следующим равенствам: 1) совместимому (между особями L+ и L+,L— и L—, D+ и D+, D— и D —), 2) зеркальному (между особями L+ и D+, L— и D—); 3) совместимому антиравенству (между особями L— и L+, D— и D+); 4) зеркальному антиравенству (между особями L+ и D—, L— и D+)- Других случаев не существует. Заметим, что первые два равенства охватываются классической теорией симметрии, а все четыре — теорией антисимметрии.
Цветную симметрию выявляют биокристаллы, побеги растений с изменяющимися по ходу стебля формами листьев, венчики цветков растений с морфологически различными лепестками и вообще все такие биообразования, качества которых могут принимать три и более различных состояний одной природы.
Симметрия подобия реализуется на биообъектах при их подобном росте и воспроизведении; она прекрасно видна на головках подсолнечника, раковинах некоторых моллюсков, верхней части побегов ряда растений.
Гомологическую, или аффинную, симметрию выявляют динамическая симметрия биокристаллов, некоторые так называемые аффинно уродливые организмы.
Криволинейную симметрию обнаруживают кроме рядов развития раковин брахиопод и цефалопод искривленные побеги стебли, корни, листья растений. Рассмотрим один из примеров подробнее. Нередко можно наблюдать, как билатерально-симметричные S-листья-(первого яруса) фасоли по мере роста искривляются, приобретая L или D конфигурацию. Мы экспериментально показали, (неопубликованные данные), что превращение S-листьев в L или D вызвано повышением содержания в меньших половинках L и D листьев ингибиторов (в частности, фенольной природы) и понижением содержания активаторов роста (типа ауксинов) и, наоборот, с повышением содержания в больших половинках этих листьев активаторов и понижением содержания ингибиторов роста. С этой картиной хорошо коррелировала и активность соответствующих ферментов и их ингибиторов. В результате, уже искусственно изменяя содержание ингибиторов или активаторов роста, например нанося их (после выделения из листьев) на те или иные половинки листа, нам удалось вызвать все мыслимые превращения форм листьев друг в друга.
Приведенные факты интересны с трех точек зрения.
Во-первых, с ботанической. Любой ботаник сказал бы, что S-лист симметричен, а L и D — асимметричны. И это было бы так с точки зрения классического учения о симметрии и совершенно несправедливо с точки зрения учения о криволинейной симметрии. Действительно, после превращения из-за неравномерного роста половинок S-листа в L или D бывшая у S-листьев прямая плоскость симметрии не исчезает бесследно, а превращается в криволинейную плоскость отражения. В результате, как и S-листья, L и D листья также по-своему зеркально-симметричны:
под действием отражения в сферическом зеркале у каждого из них меньшая половинка становится большей, большая — меньшей, а лист в целом самосовмещается.
Во-вторых, эти факты интересны "с точки зрения теории симметрии. Вплоть до последнего времени теоретики считают, что наличие в объекте зеркальных элементов исключает какую бы то ни было возможность быть этому объекту L или D. Действительно, наличие зеркальной плоскости исключает способность S-листа быть L или D, но не мешает быть L или D искривленным листьям! И это, конечно, не случайно:у S-листа зеркальная плоскость прямолинейная, сохраняющая при отражениях расстояния между соответственными точками половинок, а у L и D листьев эта плоскость криволинейная, при отражениях не сохраняющая расстояний между соответственными точками половинок, «делая» их L или D. Разумеется, сказанное верно не только по отношению к листьям, но н по отношению к любым аналогичным объектам (например, к искривленным кристаллам кварца и серы). Таким образом, ограниченно справедливым оказывается одно из самых, казалось бы, незыблемых утверждений теории структурной симметрии.
В-третьнх, эти факты интересны с точки зрения метода кристаллохимического анализа Е. С. Федорова, позволяющего по величине углов между гранями кристалла предсказывать с определенной вероятностью вещества, его слагающие. Приведенные выше биологические факты с S, L, D листьями интересны тем, что они указывают на явную возможность расширения границ федоровского метода, распространения его на биологические образооания. Можно и по их форме судить с определенной вероятностью о биофизикохимнческих и физиологических их особенностях (и наоборот). В данном случае это выразилось в том, что мы: 1) констатировали возникновение из S-листьев искривленных L и D с неравными половинками;
2) возложили «ответственность» за правизну и левизну, а также неодинаковость половинок на регуляторы роста, их ферментные системы и ингибиторы; 3) в соответствии с истинной симметрией форм S, L, D листьев построили гипотезу о пространственном распределении регуляторов роста, ферментов, ингибиторов, ожидая вполне определенные с точки зрения закономерностей форм S, L, D листьев результаты; 4) подтвердили гипотезу биохимическими анализами;
5) зная эти закономерности, по строгому плану изменили формы одних листьев в формы других.
В заключение отметим: мы не думаем, чтобы теоретико-групповое изучение биообъектов свелось к формулировке получаемых результатов на языке только уже известных теорий симметрии. Дело в том, что так или иначе выявление видов симметрии конкретных биообъектов связывается с выявлением способов упаковки тех или иных компонентов в эти биообъекты. Часть из них удавалось и наверняка удастся расшифровать на основе стандартных экспериментальных методов и методов уже известных теорий структурной симметрии. Однако для расшифровки другой части биоупаковок рамки существующих теорий структурной симметрии придется существенно расширить хотя бы для математического анализа и вывода всех возможных способов заполнения пространств без и (или) с промежутками, нежесткими и (или) жесткими, растущими и (или) нерастущими, часто неправильной конфигурации выпуклыми и (или) вогнутыми компонентами. Для лучшего уяснения этой идеи полезно сравнить способы заполнения пространства в блоках кирпичами со способами заполнения пространства в апельсинах сочными ячейками. Понятно, что выявление видов биологических упаковок поможет глубже понять сущность жизни. С другой стороны, оно может буквально революционизировать производство тары и упаковок, производство, без которого, как известно, не обходится ни одна отрасль народного хозяйства.
Похожие работы


l Какие виды симметрий, встречаются в природе; l Как применяет красоту симметрии в своих творениях человек? Поэтому тему своего исследования я назвала «Симметрия — символ красоты, гармонии и совершенства». §2. Что такое симметрия? Ее виды в геометрии. О, симметрия! Гимн тебе пою! Тебя повсюду в мире узнаю. Ты в Эйфелевой башне, в малой мошке, Ты в елочке, что у лесной дорожки. С ...

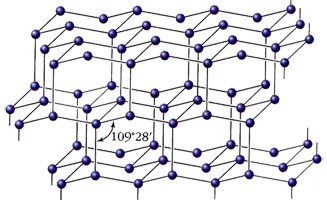
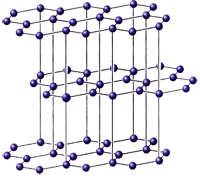
... к старинным геохимическим циклам планеты). Осуществление завершающего звена, по В.И. Вернадскому, должно быть итогом переработки отходов и вышедшей из употребления продукции автотрофными, так как высшие формы живой материи - гетеротрофные - способны усваивать без ущерба для себя лишь химически чистые, однородные элементы. Хлорофильные растения и окисляющие бактерии должны поэтому выполнить роль " ...


... требуется оценить отклонение от нулевого положения, например на руле грузовика или на штурвале корабля. Симметрия проявляется в многообразных структурах и явлениях неорганического мира и живой природы. В мир неживой природы очарование симметрии вносят кристаллы. Каждая снежинка- это маленький кристалл замерзшей воды. Форма снежинок может быть очень разнообразной, но все они обладают симметрией - ...
... по-видимому, прибавлялась только как искусственная роскошь к довольно узкому готовому миру вещей с их свойствами и силовыми взаимодействиями, их движениями и изменениями». Об определении категорий симметрии и асимметрии В настоящее время в науке преобладают определения указанных категорий на основе перечисления их важнейших признаков. Например, симметрия определяется как совокупность ...
0 комментариев