Количественное распределение жизни
Бентос
Вертикальное распределение жизни
Бентос
Батиаль
Батипелагиаль
Сравнительная оценка биомассы
Жизнь Атлантического океана и его биологические ресурсы, особенности водных экосистем
Биогеоценотические особенности Атлантического океана
Фарерско -Исландский порог и воды Гренландии
Ньюфаулендское мелководье, район Лабрадора и шельфы новой Шотландии и Новой Англии (Северо-западный сектор Атлантики)
Шельф северо-западной Африки
Саргассово море
Карибское море и Мексиканский залив
Гольфстрим
Шельф Южной Америки
Антарктическая область
Биологические ресурсы и освоение их человеком
Экологические проблемы океана (заключение)
Навигация
Ньюфаулендское мелководье, район Лабрадора и шельфы новой Шотландии и Новой Англии (Северо-западный сектор Атлантики)
Атлантический океан: биогеоценоз и экологические проблемы
200791
знак
7
таблиц
0
изображений
9.3. Ньюфаулендское мелководье, район Лабрадора и шельфы новой Шотландии и Новой Англии (Северо-западный сектор Атлантики).
Районы, входящие в северо-западный сектор Атлантики, существенно различаются между собой: шельф Лабрадора находится под воздействием холодного Лабрадорского течения, простирающегося от Баффиновой Земли; большую часть года здесь можно встретить плавучие льды и айсберги. Залив Святого Лаврентия на небольшой площади подвержен влиянию опреснения одноименной реки; шельфы Новой Шотландии и Новой Англии расположены в непосредственной близости от Гольфстрима. И, тем не менее, эти районы имеют много общего. В результате проникновения холодных лабрадорских вод на юг через проливы Бейлл-Айл и Кабота, а также вокруг Ньюфаундлендского мелководья по его склону вплоть до мыса Код, отепляющего влияния атлантических вод, заходящих в пролив Девиса и омывающих свал Лабрадорского шельфа, термический режим, придонных слоев самых северных и самых южных районов различается не так уж сильно. По свалу шельфа Лабрадора на глубине 400—500 м мы встречаем в течение всего года температуру 3,5—4,0° С; вблизи мыса Код, на западном склоне банки Джорджес, даже в августе можно наблюдать температуру менее 5° С. В результате нерест трески, главной промысловой; рыбы, происходит и на севере, на широте залива Унгава, и в заливе Мэн, вблизи мыса Код.
Основные рыбы данного района — треска, пикша, серебристый хек, морской окунь и морская сельдь — распространены в водах шельфа или его свалов при диапазоне температур не выше 8—10° С. Ни один из этих видов ни в период нереста, ни в период нагула не встречается в водах Гольфстрима. Средой обитания промысловых рыб области являются водные массы полярного и атлантического происхождения, а также воды склонов шельфа.
Резко выраженные фронтальные зоны встречаются не только на Ньюфаундлендском мелководье, но и в пределах шельфа Новой Шотландии, а также на банке Джорджес. Перепады температуры поверхностного слоя в 3—5° на протяжении нескольких кабельтовых — явление обычное. Западный сектор Атлантики — район чрезвычайно резких в первую очередь термических градиентов, которые определяют смену фаунистических комплексов. На южных склонах Ньюфаундлендского мелководья размножается мойва, а в нескольких десятках миль к югу встречаются тунцы и меч-рыба. Быстрая смена термических условий часто является причиной массовой гибели трески, пикши, мойвы, сельди в районе Ньюфаундлендского мелководья подробно описанной Темплеманом. Воды Ньюфаундлендского мелководья характеризуются высоким содержанием фосфора и кремния и обилием планктона (33,60). Ньюфаундлендское мелководье как район богатых рыбных промыслов известен человечеству более 500 лет, однако, интенсивная эксплуатация района началась только в последние 20— 25 лет.
Резкое увеличение уловов в западных районах Атлантического океана началось в 50-х годах, когда их стали посещать флотилии многих европейских стран. Этот процесс относится к тому моменту, когда в морях Европейского Севера были использованы благоприятные последствия прекращения промысла в военные годы и производительность лова в традиционных районах европейских стран начала снижаться.
Говоря об изученности этого района, следует отметить, что наряду с удовлетворительными знаниями гидрологии района до сего времени недостаточны сведения о первичной продукции, зоопланктоне, бентосе, а также о питании рыб для получения правильного представления о трофических системах и взаимосвязях. До настоящего времени нет достаточных данных о биологии рыб, имеющих важное значение в питании хищных рыб, в частности мойвы и песчанки. Мы располагаем сведениями о распределении главных промысловых рыб, но лишь несколько лет назад были обнаружены нерестилища их и изучен дрейфы молоди.
Главной причиной медленного развертывания океанографических и научно-промысловых работ в западном секторе Атлантики явилась высокая продуктивность района, обеспечивающая хорошие уловы, несмотря на минимальные знания о сырьевой базе. Промысел Канады и США базировался в прибрежных районах, что также не способствовало расширению морских исследований. Однако в последние несколько лет рыболовные лаборатории Совета по изучению рыбных ресурсов Канады сделали довольно много в изучении биологии трески, пикши и морского окуня. Существенный вклад в изучение района внесли советские исследователи, выявившие возможности промысла морского окуня на банке Флемиш-Кап, сельди и серебристого хека на банке Джорджес. Принципиально новые представления о районе в целом и взаимосвязи Лабрадорского района и Ньюфаундлендского мелководья были получены в результате исследований сотрудников ПИНРО В. П. Серебрякова (1962 г.) и А. И. Постолакня (1962, 1966 гг.), выявивших нерестилища трески на больших глубинах в северном Лабрадорском районе.
Исследователи Полярного института, хорошо представляя значение дрейфа молоди в жизни трески Баренцева моря, правильно организовали изучение миграций трески в западном секторе Атлантики и подвергли исследованиям глубинные районы Северного Лабрадора, куда, несмотря на высокие широты, проникают относительно теплые атлантические воды.
В, П. Серебряков проследил за дрейфом икринок, личинок и мальков трески от Северного Лабрадора до Ньюфаундлендского мелководья, В южной части Ньюфаундлендского мелководья значительного дрейфа молоди нет.
В результате мечения трески, проведенного А. И. Постолакием (1966 г.) на местах нереста у Северного Лабрадора, прослежено движение стай трески на нагул в юго-восточном направлении на расстоянии 800—900 км. Скорость миграции трески от Северного Лабрадора до южного района по течению составляет около 35 км в сутки, далее на юг она уменьшается до 10—20 км в сутки. А. И. Постолакий показал, что особенно активно нагульная миграция протекает у крупных рыб, которые движутся впереди стада с большей скоростью и достигают северных и восточных бухт о. Ньюфаундленд. Как предполагает большие глубины начинается в сентябре. В январе начинается движение в районы нереста.
В других районах столь значительных миграций трески, так же как и пассивного расселения икры, личинок и молоди на большие расстояния, нет.
А. И. Постолакий, уход трески из прибрежных районов на большие глубины начинается в сентябре. В январе начинается движение в районы нереста.
В других районах столь значительных миграций трески, также как и пассивного расселения икры, личинок и молоди на большие расстояния, нет.
В пределах Ньюфаундлендского мелководья треска наиболее интенсивно питается мойвой и песчанкой, второстепенное значение имеют беспозвоночные; на склонах мелководья в питании трески важную роль играют полуглубоководныс рыбы—паралепис (Paralepididae), светящиеся анчоусы (Myctophidae) и нектонные ракообразные; на банке Флемиш-Кап, как и на ее склонах преобладают полуглубоководные рыбы, встречается также молодь морского окуня; на шельфе Новой Шотландии в питании отсутствует мойва, часто попадаются мелкие экземпляры серебристого хека и урофицис.
Важнейшим компонентом питания трески Баренцева моря и Ньюфаундлендского мелководья служит мойва. Хорошим дополнением в районе Ньюфаундленда является песчанка, в Баренцевом море не меньшее значение имеют мелкая сельдь и сайка, а также эуфаузииды. Полуглубоководные рыбы в Баренцевом море в состав питания трески не входят. Они являются дополнительной кормовой базой трески в глубоководных районах Ньюфаундленда. В том и другом районе беспозвоночные играют роль дополнительного питания трески, то же, по-видимому, можно сказать о ее собственной молоди, которая периодически используется взрослой треской.
Темп роста трески Баренцева моря значительно выше темпа роста трески Лабрадора и северной части Ньюфаундлендского мелководья. Треска указанных районов самая мелкая из всех стад атлантического вида, отличается наименьшим ростом и особенно ранним созреванием (в возрасте 5—-6 лет) при длине тела около 50 см, в то время как треска Баренцева моря, Исландии и южных участков Ныофаундлендского мелководья созревает в возрасте 8—9 лет и более при длине 65—70 см.
Многочисленна в северо-западном секторе Атлантики пикша. По сравнению с треской она менее стенобионтная. Это выражается в меньшем ареале ее, колебании численности отдельных поколений, периодическом расширении и сужении ареала и изменении плотности популяции.
Серебристый хек (Merluccius bilinearis) относится к семейству мерлуз, широко распространенных в северном и южном полушариях Атлантического и Тихого океанов. Данный вид встречается только в Атлантическом океане, у берегов Америки, между 45 и 37° с. ш. Серебристый хек относится к мелким мерлузам — длиной до 45, редко 50 см. Интенсивный нерест его происходит по южному склону банки Джорджес и в районе банки Сейбл, находящейся в северной половине Новошотландского шельфа. Половозрелые особи и развивающаяся икра были найдены также в юго-западной части Ньюфаундлендского мелководья.
Нерест происходит с июня по конец августа. Выклюнувшиеся личинки дрейфуют в соответствии со схемой течений в южном направлении на расстояние 200—300 км. В первые годы жизни серебристый хек питается планктонными ракообразными, из которых главное значение имеют Thysa-noessa inermis, Thysanoessa raschii, Meganicti phanes norvegica,. Thernisto compressa. Особи крупнее 30—32 см питаются рыбой — мелкой сельдью, аргентиной, песчанкой, а также молодью кальмаров и креветками.
В 1960 г. Международная комиссия по рыболовству в северо-западной Атлантике и Международный совет по изучению моря (ИКЕС) организовали в Копенгагене симпозиум по морскому окуню, в котором приняли участие ученые большинства стран заинтересованных в эксплуатации его запасов.
В западном секторе Атлантики встречается глубоководный вид Sebastes rnentella Travin, обитающий на склонах шельфа от южных границ Лабрадора до мыса Код на глубинах до 600— 700 м. Таким образом, ареал его значителен по широте, простираясь от 55 до 40° с. ш., но ограничен по площади. Вымет личинок происходит на большей части свала шельфа, где встречается окунь. Это обстоятельство говорит о том, что морского окуня нельзя отнести к активным мигрантам, у которых нерестилища обычно обособлены от районов нагула.
Высказывались предположения о том, что морские окуни, и в частности Sebastes mentella, живут в толще воды и вблизи дна встречаются только на глубине горизонта их скоплении. Естественно, что при таком образе жизни скопления морского окуня на склонах шельфа всегда пополнялись бы за счет «глубинного пелагического запаса» и запасы его на шельфе были: бы стабильны и не уменьшались под воздействием промысла. Миграции окуня не могли быть изучены путем мечения, так как большинство особей, поднятых на палубу с глубин, нежизнеспособны из-за кессонных явлений. Большие трудности вызывало определение возраста у окуня, но их удалось преодолеть. Темп роста морских окуней оказался исключительно низким. Только впервые несколько лет он увеличивается за год на 3—5 см, затем годовые приросты его не превышают 1,0—1,5 сантиметров. Половая зрелость у морского окуня наступает не раньше 12—15 лет, т. е. в возрасте, в 3 раза большем, чем возраст созревания большинства морских сельдей, и почти в 2 раза большем, чем - возраст созревания трески.
Чрезвычайно низкий темп роста морских окуней и позднее наступление у них половой зрелости вполне объясняют причины снижения их уловов под воздействием чрезмерно интенсивного рыбного промысла.
Ареал сельди по западному побережью Атлантического океана ограничен заливом Мэн, банками Джорджес и Банкеро, заливами Св, Лаврентия и Фанди-Бай; встречается сельдь и у океанского побережья о. Ньюфаундленд.
Весьма вероятно, что сельди банки Джорджес, заливов Мен и Фанди представляют единое стадо. На банке Джорджес происходит размножение и нагул подрастающих рыб. Молодь этого стада первые годы жизни проводит в прибрежных районах заливов Мэн и Фанди. Большая часть половозрелого стада находится на байке Джорджес круглый год. Средой ее обитания являются водные массы шельфа и зона конвергенции их с водными массами свала. По данным Е. А. Павштикс, эта область весьма продуктивна и биомасса зоопланктона, в основном калянуса, достигает 500 мг/м3.
В 1964 г. АтлантНИРО по количеству отложений икры определен примерный запас взрослых сельдей на банке Джорджес, равный 1200 тыс. т. Следовательно, общий запас, включая неполовозрелую часть популяции, может составить около 1800 тыс. т.
В западном секторе Атлантики живет ньюфаундлендское стадо гренландского тюленя (Phobia groenlandica), биология которого связана с районом Лабрадора, прилегающими участками Ньюфаундлендского мелководья и заливом Св. Лаврентия. В первой половине XIX в. годовая добыча тюленя составляла в среднем около 500 тыс. голов. В дальнейшем добыча его сокращалась и в последнем десятилетии прошлого века составила в среднем около 240 тыс. голов в год. В течение 1949—1958 гг. общая добыча повысилась до 320 тыс. голов, включая район западного побережья Гренландии. По результатам аэрофотосъемок в 50-х годах запас ньюфаундлендского тюленя был определен в 3,3 млн. голов.
Надо полагать, что основным объектом питания тюленей должна быть мойва, ареал которой совпадает с ареалом тюленя и которая является главной пищей его во всех районах атлантического сектора.
В прибрежных водах Канады встречается омар, пользующийся большим опросом на рынках Канады и США.
9.4.Шельф Западной Европы.
В шельф Западной Европы входят моря: Северное, Балтийское, Ирландское, Датские проливы, Ла-Манш.
Рассматриваемый район подвержен влиянию теплых атлантических вод, однако, к востоку это влияние ослабевает. Высокая соленость, приближающаяся к атлантической, сохраняется только в Северном море (33—34°/оо). Балтийское море должно быть отнесено к солоновато водным бассейнам (5—6°/оо). Морской климат островов Великобритании и Северного моря сменяется на более континентальный Балтийского моря.
Состав ихтиофауны с запада на восток резко сокращается. У берегов Ирландии встречаются все виды рыб Северной Атлантики, в Северном море число видов важных промысловых рыб уменьшается до 10—12, в Балтийском море их всего два— треска и салака. Третий объект — шпрот — является скорее специфическим видом Балтийского моря и Датских проливов. Главные рыбы шельфа Западной Европы — треска и сельдь— образуют ряд популяций (стад), отличающихся размерами, темпом роста, возрастом наступления половой зрелости и сроками размножения. Среди сельдей в этом районе выделяются две экологические группы, размножающиеся весной в солоноватых водах, вблизи рек, и при высокой солености — вдали от берегов, на банках. Размножение их происходит летом и осенью. Численность первой группы сельди крайне низка в Северном море и достигает максимума в Балтийском; вторая группа, наоборот, многочисленна в Северном море и малочисленна в Балтийском.
В Английский канал и Северное море из более южных районов заходят скумбрия, сардина, ставрида и анчоус.
Промысел сельди возник в Северном море очень давно и сыграл важную роль в экономике прибрежных стран. Однако до начала XX в., несмотря на большую плотность населения окружающих берегов, уловы в Северном море были небольшими. В конце прошлого века общий улов в рассматриваемом районе не превышал, по-видимому, 1 млн. т.
Северное море даст почти 70 % всей продукции района, за ним следуют Балтийское море и Датские проливы, Ирландское морс и воды, омывающие острова Великобритании.
Площадь Северного моря составляет 544 тыс. км2 средняя глубина—96 м. Объем равен 42 тыс. км3. Основной водообмен происходит с севера, между Шетландскими островами и Норвегией. Приход здесь составляет 25 тыс. км3, расход — 26,8 тыс. км3. Приход через Дуврский пролив равен всего 1,8 тыс. км3, расход — 0,8 тыс. км3. Незначителен обмен и с Балтийским морем: приход равен 1,7 тыс. км3 и расход — 1,2 тыс. км3. Водообмен с соседними районами по отношению к объему моря несколько превышает 55%. Речной сток (0,3 тыс. км3) составляет около 0,6%. Эти цифры говорят о том, что биогенные элементы материкового происхождения не могут иметь серьезного значения для водоема в целом, хотя вблизи речных устьев их роль может быть значительной.
Процессы биологического продуцирования в Северном море носят сезонный характер, что связано с изменением солнечной радиации, температуры воды, а также с характером вертикальной циркуляции. Начало весенних процессов в южных районах моря наблюдается с конца февраля, в северных—с апреля. Продолжительность вегетационного периода в мелководных юго-восточных районах составляет около 7 месяцев, на северо-западе — свыше 9 месяцев. Среднее количество фосфора к весне достигает 25 мг/л. Первичная продукция, по Стиллу, для северных районов составляет 80 гСм2/год. Биомасса планктона в мае—июне равна 200—500 мг/м3. Бентос изучен плохо. Биоценозы не установлены. Биомасса бентоса в северных районах моря не превышает нескольких десятков граммов на 1 м2 («Гидрометеорологический справочник Северного моря», 1964).
К планктоноядным рыбам, населяющим Северное море, должны быть отнесены сельдь, песчанка, тресочка Эсмарка (Trisopterus esmarki); к бентосоядным—камбалы, скаты, по-видимому, мерланг и пикша, однако в питании последней значительную долю составляет икра сельди, т. е. продукция планктона. Смешанным питанием характеризуется треска и хищным — сайда.
Биология морской камбалы, пикши и сельди, главных объектов промысла в Северном море, интенсивность использования которых достигло предельной величины, изучена особенно подробно.
Воздействие интенсивного промысла в Северном море, прежде всего, сказалось на морской камбале благодаря ее донному образу жизни, малой подвижности, а также высокой стоимости, которая исключала возможность экономического регулирования ее добычи. В 30-х годах вследствие технического усовершенствования тралов и увеличения усилий промысла воздействие его стало сказываться и на пикше. Влияние промысла на запасы сельди стало очевидным после второй мировой войны, когда в итоге прекращения промысла в военные годы увеличились уловы на усилие, и повысился возрастной состав.
Эксплуатация запаса морской камбалы показала, что интенсивность лова приводит к изменениям возрастного состава. Вслед за усилением промысла возрастной состав снижается, при ослаблении рыболовства увеличивается численность старших возрастов и средний возраст. Средний возраст морской камбалы изменялся в пределах 3,4—5,3 года.
Северное море можно рассматривать в какой-то мере центром ареала пикши (Melanogrammus aeglefinus) в северо-восточном секторе Атлантики. Здесь по сравнению с Баренцевым морем и районом Исландии она более многочисленна и менее подвержена воздействию климатических условий.
Пикша размножается в северной части моря. Южная граница нерестилищ пикши тянется до пролива Скагеррак. Придонные температуры в районах размножения пикши сравнительно высоки (6—8° С), так же как и соленость (свыше 35°/оо). Нерестовые миграции молодых рыб ограничены и совершаются между хорошо прогреваемыми мелководьями и более глубокими участками моря. С возрастом миграции пикши расширяются к северу и востоку.
Выклюнувшиеся из икринок личинки пикши ведут пелагический образ жизни и, прежде чем к концу лета перейти к донному образу жизни, расселяются на широком пространстве Северного моря. Явление флюктуаций пикши в Северном море выражено менее резко, чем в Баренцевом море, однако и здесь наблюдаются богатые и бедные поколения ее. Многочисленные поколения пикши отличаются замедленным темпом роста, что ранее связывали с влиянием пищевой конкуренции предшествующих богатых поколений. Однако уже в 30-х годах было установлено преобладающее влияние развивающегося промысла не только на возрастную структуру популяции, что сказалось не только на устойчивом уменьшении старших возрастных групп пикши, но и на возросшем темпе роста молодых рыб.
Большой интерес представляют взгляды Райта на воспроизводство пикши при резком изменении возрастной структуры ее стада. В возрасте двух лет пикша средних размеров выметывает 30 тыс. икринок. В возрасте от двух до трех лет она удваивает свою массу и утраивает количество вымотанных икринок. В возрасте от двух до четырех лет пикша утраивает свою массу и увеличивает количество икринок в 5 раз. Между вторым и пятым годом жизни пикши масса ее увеличивается почти в 4 раза, а количество икринок — в 7 раз. Наконец, между двумя и шестью годами жизни пикши масса ее увеличивается в 5 раз, а плодовитость — в 9 раз. Одна шестигодовалая самка пикши массой 596 г выметывает 278 тыс. икринок.
Райт отмечает, что, вылавливая основную массу производителей на четвертом году жизни, мы там самым лишаем район большей части его производителей и принуждаем двухлетних и трехлетних рыб, мало приспособленных природой для воспроизводства, принять его на себя.
Сельди (Clupea harengus harengus) Северного моря — объект интенсивных исследований в течение почти уже 70 лет. Тем не менее, многие вопросы их жизненного цикла остаются спорными. И, прежде всего мы не располагаем вполне достоверными данными о распределений и миграционных контурах отдельных популяций, которые, несомненно, имеются в Северном море, обладают спецификой и оказывают важное влияние на сезонный ход промысла и эксплуатацию их запаса. Следует отметить, что все расовые исследования, проводившиеся в течение многих десятилетий учеными различных стран, дали для понимания этого вопроса гораздо меньше, чем мечение сельдей, осуществленное в последние 10—15 лет. А также изучение результатов промысла сельди в отдельных районах, предпринятое Международным советом по изучению моря в связи с возросшей интенсивностью сельдяного рыболовства («Herring tagging...», 1954; Gushing, 1967; Parrish, 1962; Parrish and Craig, 1957; Parrish and Seville, 1965).
В результате длительного обсуждения было принято следующее разделение сельдей, размножающихся в северо-западной части Северного моря, вблизи берегов Шотландии. Места их откорма находятся северо-восточнее районов размножения, а зимовка проходит где-то вблизи Норвежского желоба. Две нерестующие группы сельдей на банке Доггер, откармливающихся в северных районах моря, зимуют вблизи южной оконечности Скандинавского полуострова; сельди Даунс, размножающиеся в Ла-Манше, в частности на банке Сандети, зимуют в центральных частях Северного моря, вблизи банки Доггер.
Следует отметить ряд общих черт, отличающих сельдей Северного моря от других рас атлантического подвида. Жизненный цикл и миграционные контуры их ограничиваются Северным морем. Они мельче атлантическо-скандинавской расы нередко достигают длины 30 см. Половозрелыми становятся преимущественно в 4—5, изредка в 3—6 лет. Размножаются летом и осенью, причем в северных районах раньше, чем в южных. Откорм и рост совершаются в два этапа — до размножения и после. В размножении участвуют три-четыре возрастные группы. Пополнение запаса относительно других рас сельдей, и в частности атлантическо-скандинавских, стабильное. Явление флюктуации выражено слабо. Основу питания составляет зоопланктон, по-видимому, существенное значение имеют эуфаузииды. Врагов у сельдей Северного моря немного. Во время размножения много, их икры поедает пикша. Молодую и подрастающую сельдь в больших количествах истребляет сайда и акула катран.
История развития сельдяного промысла в Северном море изучена достаточно подробно с начала столетия.
В середине 50-х годов возникает новый вид промысла сельди, получивший название индустриального рыболовства, — траловый лов молоди сельди, идущей на производство кормовой муки и жира, в юго-восточных районах моря. С этого времени лов сельди происходит по всей акватории Северного моря.
В 1957—1958 гг. этот промысел достиг расцвета: общий улов молоди превышал 150 тыс, т, составляя около 1 5 улова всей сельди в Северном море по массе и почти 2/3 по числу экземпляров. В последующие годы отмечено резкое уменьшение уловов сельди у берегов Англии и в южных районах Северного моря.
Однако общий улов взрослой и мелкой сельди не уменьшился, колеблясь от 640 до 850 тыс. тонн. Таким образом, за весь период развития сельдяного промысла в Северном море наблюдались три максимума улова: в 1913 г.—около 700 тыс. т, в 1937 г.—680 тыс. т и в 1956—1963 гг. средний улов за год— 750 тыс. т. За эти же годы средний улов молоди сельди составил 120 тыс. тонн. Таким образом, средний годовой улов взрослой пищевой сельди составил 630 тыс. т, т. е. меньше, чем в 1913 и 1937 гг.
Международному совету по изучению моря удалось подсчитать усилие промысла (количественная оценка его интенсивности) с 1946 по 1965 г. и определить улов на усилие взрослой сельди для северного, центрального и южного районов моря по траловому и дрифтерному промыслу. Использовав для этого данные об улове и сведения об усилии промысла большинства стран, участвующих в сельдяном рыболовстве в Северном море. За 1946—1963 гг. улов на усилие в траловом промысле снизился по всем районам. Для дрифтерного лова характерно максимальное снижение улова в южной части моря, меньшее— в центральных районах и некоторое повышение — в северной части. Однако в 1964 г., по данным Абердинской станции, уловы на усилие в дрифтерном промысле снизились.
Таким образом, можно констатировать, что рост интенсивности сельдяного рыболовства не привел к увеличению улова пищевой сельди, а производительность промысла даже снизилась в целом по морю более чем наполовину. Развитие лова молоди и мелкой сельди на банке Блодептрунд дало аредие годовую прибавку около 120 тыс. т, однако при этом почти на эту же величину уменьшились абсолютные уловы взрослой пищевой сельди в южной части моря.
С повышением интенсивности сельдяного рыболовства в уловах уменьшалась доля старших возрастных групп, сельди стали лучше расти и раньше созревать, но в то же время, возросшая интенсивность промысла сельди, изменив размерно-возрастную структуру запаса, не принесла желаемого результата — увеличения улова.
Происшедшие изменения в состоянии запаса камбалы, пикши и сельди Северного моря имеют важное теоретическое значение при подходе к оптимальной интенсивности рыболовства.
В результате интенсивного промысла резко уменьшился процент старших возрастных групп, темп роста их повысился, и возраст наступления половой зрелости снизился, улов на усилие упал.
Показанные изменения биологических свойств популяции говорят о том, что в условиях Северного моря, где ареалы всех трех видов рыб ограничиваются его акваторией, кормовые ресурсы моря определяют рост и планктоноядных и бентосоядных рыб. При разрежении популяции рост особей повышается в результате обеспечения их кормом. Повышение роста приводит к более раннему наступлению половой зрелости. Следовательно, кормовые ресурсы водоема способствуют более интенсивному росту молодых рыб. Количество пищи, расходуемой на поддержание старших возрастных групп рыб, должно было снизиться.
Основываясь на этих положениях, естественно было ожидать повышения продукции хозяйственно-ценных объектов водоема. Но именно этого и не наблюдалось в Северном море в отношении всех трех видов рыб при увеличении интенсивности их промысла. Это обстоятельство заслуживает самого пристального внимания. Улучшение роста при разрежении популяции и уменьшении в ее составе старших возрастных групп, несомненно, должно приводить к более эффективному использованию кормовых ресурсов водоема. Если же этого не наблюдается, то возникает мысль, что при разрежении популяции часть кормовых ресурсов водоема используется более интенсивно, а другая часть либо не используется вовсе, либо используется слабо. Трудно допустить, что воспроизводство не обеспечивало уже той численности рыб, которая гарантировала бы использование кормовых возможностей водоема. Тогда становится более вероятным недоиспользование части кормовых ресурсов при разрежении популяции и изменении ее возрастной структуры. Ответ на этот вопрос может быть получен только после тщательного изучения питания каждого вида рыб в течение всей его жизни. Одинаковая продукция сельди при различной интенсивности использования ее запасов в какой-то мере подтверждает наши предположения о том, что старшие возрастные группы рыб не являются в полной мере конкурентами более молодых особей и что проблема обеспеченности их пищей решается в результате ряда приспособлений, которыми не обладают более молодые возрастные группы (большая подвижность старших особей, возможность заглатывания более крупных кормовых организмов, откапывания их в грунте и т. д.).
Из семейства тресковых кроме пикши Северное море населяют треска, сайда, мерланг. До последнего времени, сайда считалась второстепенным объектом промысла, и улов ее был незначительным, однако уже с 50-х годов добыча сайды резко увеличилась.
Особенно значительный промысел рыбы, идущей для технического использования, развился после войны в Норвегии, где более половины улова сельди стали перерабатывать на утилизационных фабриках. В Северном море это произошло несколькими годами позже. В начале 50-х годов в юго-восточных районах Северного моря возник лов молоди сельди. В это же время организуется специальный лов песчанки и тресочки Эсмарка.
Похожие работы
... обсуждал взаимодействие организмов со средой с позиций, близких дарвиновским. Они были предвестниками эволюционной идеи и целостного восприятия природных комплексов, состоящих из живых и неживых компонентов. Большой вклад в развитие экологических представлений в этот период внесли российские естествоиспытатели А.Т. Болотов (1738 - 1833), И.И. Лепехин (1740 - 1802), П.С. Паллас (1741 - 1811). Во ...

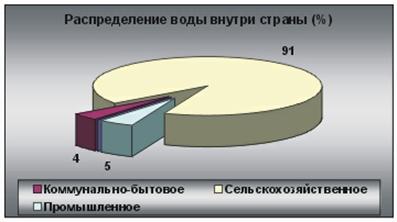
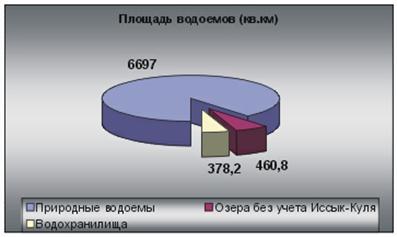
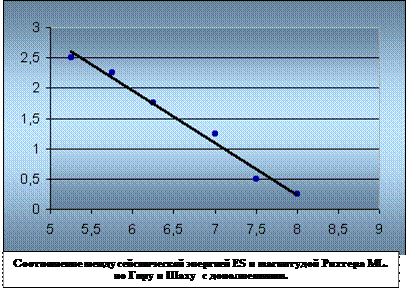

... предыдущего чрезмерного стравления, в восстановлении будут преобладать разновидности сорняков. Глава 3. Проблемы горных территорий и возможные способы их решения. Природные и антропогенные катаклизмы. Природные катаклизмы в горах представляют результат геотектонической природы гор и их экологических характеристик. Однако катаклизмы зачастую вызываются деятельностью человека. Перед ...
... обыкновенный, орлан-белохвост, сапсан, кречет, осетр сибирский, голец арктический, муксун, тугун, хариус сибирский. 3. Влияние динамики изменения площади особо охраняемых территорий на сохранение биоразнообразия 3.1 Динамика численности охотничье-промысловых животных В последние 30 лет все компоненты ландшафтов северной тайги Западной Сибири, включая орнитокомплексы, испытывают мощное ...
... , а целое – на части. Анализ и синтез. Антропный принцип. Принципы универсального эволюционизма. Вместе с представлением о коэволюции - составляющие эволюционно-синергетической парадигмы. Естествознание из ценностно-нейтрального знания превращается в аксиологически ориентированное, предполагающее введение этических, эстетических и т.п. норм в научное исследование. Биоэнергоинформатика – новое ...
0 комментариев