Количественное распределение жизни
Бентос
Вертикальное распределение жизни
Бентос
Батиаль
Батипелагиаль
Сравнительная оценка биомассы
Жизнь Атлантического океана и его биологические ресурсы, особенности водных экосистем
Биогеоценотические особенности Атлантического океана
Фарерско -Исландский порог и воды Гренландии
Ньюфаулендское мелководье, район Лабрадора и шельфы новой Шотландии и Новой Англии (Северо-западный сектор Атлантики)
Шельф северо-западной Африки
Саргассово море
Карибское море и Мексиканский залив
Гольфстрим
Шельф Южной Америки
Антарктическая область
Биологические ресурсы и освоение их человеком
Экологические проблемы океана (заключение)
Навигация
Биогеоценотические особенности Атлантического океана
Атлантический океан: биогеоценоз и экологические проблемы
200791
знак
7
таблиц
0
изображений
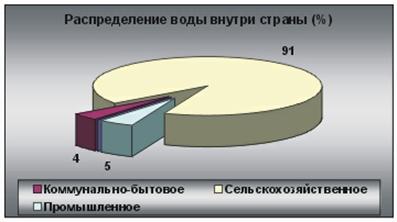
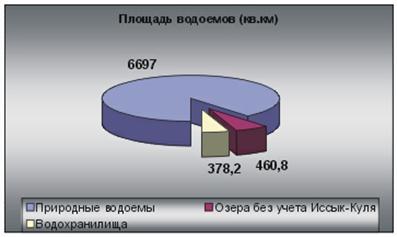
9. Биогеоценотические особенности Атлантического океана.
Атлантический океан обладает рядом геоморфологических, гидрологических и гидрохимических особенностей, определяющих его жизнь, распределение и миграции его гидробионтов, трофические системы, биопродукцию.
Из всех океанов он имеет наибольшую по меридиану протяженность—около 8,5 тыс. миль. Северной границей его считается Фарерско-Исландский порог, но воды его в поверхностной толще не только доходят до Новой Земли и Шпицбергена, но и проникают в Ледовитый океан, образуя там теплую прослойку воды, открытую еще Нансеном во время дрейфа на «Фраме».
Важное значение в гидрологии Атлантического океана играет приток полярных вод, образующий холодное Восточно-Гренландское течение, дающее начало фронтальной зоне Северной Атлантики, простирающейся далеко на юг вдоль берегов Северной Америки, и образующее ответвление к юго-востоку по направлению к Фарерским островам.
Три моря—Баренцево, Гренландское и Норвежское — общей площадью более 8,5 млн. км2, формально входящие в границы Ледовитого океана, по своей природе неразрывно связаны с Атлантическим океаном. Воды атлантического происхождения образуют в них мощный теплый приповерхностный слой, в котором протекает жизнь. Ареалы важнейших промысловых рыб — трески, пикши, сельдей, морских окуней, обыкновенного и синекорого палтусов — захватывают северные воды Атлантического океана и названные моря. Потоки атлантических вод представляют собой миграционные пути этих и многих других видов рыб и приводят их от мест размножения к местам нагула.
Все это заставляет в данной главе, где рассматриваются жизнь, трофические системы и биологические ресурсы Атлантического океана, коснуться хотя бы в краткой форме биологии этих морей, представляющих собой своеобразный биогеоценоз, отличающийся от Тихого и Ледовитого океанов и в то же время носящий в себе черты того и другого.
В жизни этого региона важную роль играют подводные хребты, пересекающие его по широте и оказывающие влияние на его гидрологический режим.
Для северной половины Атлантического океана характерен развитый шельф: на востоке — Северное и Ирландское моря, на западе — ньюфаундлендское мелководье.
Весьма значительное по площади мелководье имеется у берегов Аргентины, на нем располагаются Фолклендские (Мальвинские) острова.
Солнечная радиация, минимальная на Крайнем Севере и юге, составляет 60—65 ккал-см2/год и возрастает на экваторе до 160 ккал.
Резко отличается продолжительность дня и ночи, увеличиваясь от экватора к северу и югу, превращается у полюсов в сплошную ночь и непрерывный день в течение полугода. Для Атлантического океана характерен большой приток речных вод, сравнительно равномерно распределенный по широте: на западе — Св. Лаврентия, Миссисипи, Амазонка, Ла-Плата, на востоке — реки Балтийского бассейна, Нигер, Конго.
Важной особенностью Атлантического океана являются средиземные моря — Мексиканский залив и Карибское море на западе, и Средиземноморский бассейн на востоке, вдающийся на тысячи километров в глубь материка,
Для Атлантического океана велико разнообразие механизмов, обогащающих фотический слой биогенными элементами глубинных вод: больших по площади и значительных по активности анвеллингов у берегов Португалии, Северо-Западной и Юго-Западной Африки, конвективное перемешивание в зонах смешения атлантических вод и вод Полярного бассейна, простирающееся на юг Исландии и к Ньюфаундленду. Локальные зоны смешения наблюдаются также в Северном море, у южных берегов Норвегии, у Южной Америки вдоль Фолклендского течения. Обогащение фотического слоя биогенными элементами имеет место в ряде районов в результате обтекания неровностей береговой линии и подводных возвышенностей. Несомненную роль в обогащении афотического слоя в тропической зоне играет Экваториальное подповерхиостное противотечение Ломоносова. Наконец, известная роль принадлежит и биогенному стоку с речными водами.
Биологическая структура Атлантического океана асимметрична и по широте, и по долготе, что связано с геоморфологией чаши океана, его берегов и направленностью крупномасштабных течений.
В северном полушарии важнейший элемент гидрологического режима — Гольфстрим со всеми его разветвлениями, основной движущей силой которого являются пассатные ветры. Заслуживает внимания тот факт, что в формировании Гольфстрима играют роль оба потока Пассатного течения—полностью северный и в значительной мере южный. На юг, к берегам Бразилии, приносится теплых вод сравнительно немного.
Под воздействием проникающих через пролив Дрейка тихоокеанских вод теплый поток, идущий от экватора вдоль берегов Бразилии к югу, отклоняется к востоку, вливаясь в Великий Восточный дрейф, который представляет собой своеобразный гидрологический барьер, препятствующий меридиональным миграциям рыб, крайне характерным для северного полушария.
Не менее значительны различия западных и восточных районов Атлантического океана. На Крайний Север теплые атлантические воды проходят через восточные районы, в этих же широтах на запад идет постоянный приток полярных вод.
В бореальных широтах в западном секторе Атлантического океана течения широким фронтом омывают западное побережье Европы. В связи с такой ситуацией в западном секторе Атлантического океана наблюдаются значительно большие горизонтальные градиенты, чем на востоке.
Материал данной главы излагается по крупным районам Атлантического океана с учетом их гидрологических, гидрохимических основ, уровня биологической продуктивности, т. е. тех условий, которые определяют формирование биологических циклов и продукцию хозяйственно-ценных объектов.
Опыт эксплуатации водных биологических ресурсов планеты показал, что Атлантический океан — самый продуктивный. В то же время опережающие темпы использования его биологических ресурсов, в первую очередь в его северной части, убеждают, что скоро может наступить предельный уровень использования его рыбных богатств. Общий улов Атлантического океана в течение последних семи лет не проявляет тенденций роста, несмотря на расширение районов промысла, вовлечение новых объектов лова и повышающуюся интенсивность рыболовства в старых районах. Доля Атлантического океана в мировом улове падает: за последние 17 лет она снизилась с 46 до 39%.
Это убеждает в том, что использование биологических ресурсов океана должно быть более рациональным. Сохранение и увеличение продуктивности морского рыболовства возможно только при организации регулируемого промысла, который должен дать человечеству дополнительные белковые ресурсы и оградить океан от оскудения его живых богатств.
9.1.Северные моря и их связь с Атлантическим океаном.
Пройдя Фарерско-Исландский порог, атлантические воды устремляются к северу, образуя так называемое Норвежское течение, которое дает начало Шпицбергенскому и Нордкапскому течениям, продолжающим путь теплых вод к северу и северо-востоку.
На своем пути на север и северо-восток, атлантические зоны встречают Шпицбергенский архипелаг, острова Земли Франца-Иосифа; и Новую Землю. Благодаря этому в район к югу и западу от указанных островов ограничено проникновение холодных вод Ледовитого океана.
В то же время часть относительно теплых вод, пришедших с юга, образуя большой круговорот в Баренцевом море, направляются к юго-западу, в Норвежское море.
Основной поток холодных полярных вод спускается к югу вдоль берегов Гренландии, к мысу Фарвель. При подходе Гренландского течения к Исландии часть вод направляется к восточным берегам Исландии и далее к Фарерским островам.
Солнечная радиация северных морей невелика (около 60— 70 ккал-см2/год), и 2,5 раза меньше чем в тропиках, но т. к. эта величина приходится всего на 4—5 месяцев, то рассматриваемая зона должна быть отнесена к относительно богатой солнечной энергией. Так как в период длительной полярной ночи в итоге конвективного перемешивания вод происходит накопление биогенных элементов, то создаются весьма благоприятные условия для фотосинтеза и формирования биомассы фитопланктона, напоминая в какой-то мере исключительно быстрый рост растительного покрова в Заполярье,
Наибольшее значение среди фитопланктона имеют диатомовые, а среди зоопланктона — ракообразные (Calanus finmarchicus, Calanus hyperboreus), несколько видов эуфаузии. Биомасса зоопланктона летом превышает 500 мг/м3, достигая в отдельных случаях 1000 мг/м3.
Зона повышенной продуктивности северных районов Баренцева моря является местом нагула двух массовых, стайных, пелагических планктоноядных рыб — сайки (Boreogadus saida) и мойвы (Mallotus villosus). Совершая нерестовые миграции к южным берегам моря, они имеют исключительно важное значение в питании трески, пикши, ряда других рыб, беломорской популяции гренландского тюленя и многих видов морских птиц, гнездящихся на Новой Земле и побережье Мурмана.
Довольно обыден в северных районах Баренцева моря вблизи кромки плавучих льдов белый медведь (Ursus maritimus), .привлекаемый скоплениями сайки и мойвы, а также присутствием тюленей.
Наиболее продуктивными областями Баренцева моря являются его южная часть, до 73° с. ш., и западная, до 35° в. д. Температура атлантических вод, проникающих в Баренцево море, летом составляет 8—12° С, зимой 4—5° С, соленость — около 35%. Все весенние процессы развития жизни в море начинаются с юго-запада, от берегов Норвегии, где особенно рано начинается «цветение», а за ним размножение калянуса. С теплыми водами Атлантики в Баренцево море приносятся вместе с пищей мальки всех многочисленных промысловых рыб: трески, пикши, сайды, сельди, морского окуня, размножающихся у северо-западных берегов Норвегии. Непосредственно в Баренцевом море размножаются, как мы уже отмечали, лишь мойва и сайка, а также камбалы, зубатки; для остальных видов рыб Баренцево море только район нагула.
Значение пассивного расселения молоди промысловых рыб Баренцева моря трудно переоценить. Оно способствует очень быстрой реакции отдельных видов рыб на изменяющиеся условия водоема. Ареал молоди пульсирует от года к году и в течение года в зависимости от термического режима моря, связанного с напором атлантических вод и скоростью движения водных масс в пределах отдельных ветвей Нордкапского течения.
Молодь расселяется с водами, в которых она появилась на свет и которые медленно трансформируются на своем пути к востоку и северу. Вместе с молодью распространяется ее пища в виде зоопланктона, основные формы которого в течение лета увеличиваются в размерах. С наступлением гомотермии молодь оседает на дно и переходит на питание главным образом эуфаузиидами, распространение которых приурочено к водам атлантического происхождения.
На втором-третьем году жизни молодь трески, сельди начинает откочевывать к западу, концентрируясь вблизи ветвей Нордкапского течения. Летом ареал подрастающих рыб расширяется к востоку и северу, а зимой смещается к западу. С каждым годом они все дальше и дальше уходят в западные районы моря. К моменту полового созревания рыбы зимуют уже в непосредственной близости от нерестилищ и с наступлением весны приходят к местам размножения.
Взрослые рыбы в период между нерестами совершают этот миграционный цикл за год. Наибольшие массы атлантической воды устремляются на север через Норвежское и Гренландское моря. В районе пролива Фрама часть вод поворачивает к югу, часть переливается в Северный Ледовитый океан.
С потоками атлантической воды проникает на север через Норвежское и Гренландское моря обильная и разнообразная жизнь. Основная масса планктонных организмов перезимовывает на глубине. Нерест калянуса и эуфаузии совпадает со вспышкой развития водорослей феоцистис - началом биологической весны. Незадолго до массового развития водорослей калянус и эуфаузииды образуют преднерестовые скопления у поверхности биомасса быстро возрастает до 500 мг/м3. После нереста в поверхностном слое появляются личиночные стадии зоопланктона, которые быстро растут, увеличивая биомассу планктона до 800—1000 мг/м3 .
В южной части Норвежского моря биологическая весна наступает в марте-апреле, в северной - в мае. В июне весна доходит до Медвежинской банки, а в июле она достигает уже Западного Шпицбергена. Развитие планктона наблюдается не только в северном направлении, но и в западном, по направлению к фронтальной зоне. С теплыми водами, с развивающимся зоопланктоном дрейфуют личинки и мальки трески, сельди, морских окуней, путассу. Одновременно наблюдается миграция на север отнерестовавшейся трески и сельди. В это время в трофических системах наблюдаются весьма люболытные явления. Основа пищи - сельди —планктон, но нередко в желудках ее можно встретить молодь кальмаров и мальков, наиболее важный хищник сельдей - треска в первые месяцы жизни сама представляет собой жертву сельди. Часто в конце лета в желудках крупной сельди можно встретить
В Норвежском море широко распространена планктоноядная тресочка— Micromesistius poutassou. Размножение трески, пик сельди наблюдается на большом протяжении Норвежского моря, но значительный нерест трески происходит в Вест фьорде и на прилегающем шельфе. Главные нерестилища сельди находятся у юго-западного побережья Норвегии, к северу и югу от Бергена, а также у северо-западных берегов, на склоне мелководий между островами Рест и Сере.
В центральных областях Норвежского моря сравнительно часто встречаются молодые лососи. Летом в атлантические воды приходит скумбрия. В теплые годы она иногда посещает и побережье Сурмана. Акулы в открытой части Норвежского моря и в его восточной части встречаются исключительно редко. За многие годы работы в этом бассейне автору довелось видеть всего 2 - 3 раза сельдевых акул (Larnna cornubica), запутавшихся в сельдяных сетях. Полярная акула (Soinniosus microcephalus), сравнительно часто попадающая в тралы при лове трески в Баренцевом море, ни разу не была встречена автором этих строк в Норвежском море, в районах сельдяного промысла,
Область стыка теплых атлантических и холодных полярных вод когда-то изобиловала китами. История сохранила сведения о китовом промысле, который существовал здесь с конца XVI в. до середины прошлого века.
Сейчас киты в Норвежском и Гренландском морях крайне редки. Несмотря на полный запрет их добычи, запасы их не восстановилось. Вблизи о. Медвежий сравнительно часто наблюдаются бутылконосы (Hypereodon rostratus). Косатки (Oica orca) встречаются, но редко, еще реже можно встретить кашалотов (Physeter catodon). В южных районах, ближе к Исландии и Фарерским островам, можно увидеть небольшие стаи гринд (Globicephalus melas). Гренландский тюлень (Phoca groenlandica) и хохлач (Cystophora cristata) распространены обычно в западном секторе Гренландского моря, вблизи полярных льдов и в разводьях.
Похожие работы
... обсуждал взаимодействие организмов со средой с позиций, близких дарвиновским. Они были предвестниками эволюционной идеи и целостного восприятия природных комплексов, состоящих из живых и неживых компонентов. Большой вклад в развитие экологических представлений в этот период внесли российские естествоиспытатели А.Т. Болотов (1738 - 1833), И.И. Лепехин (1740 - 1802), П.С. Паллас (1741 - 1811). Во ...


... предыдущего чрезмерного стравления, в восстановлении будут преобладать разновидности сорняков. Глава 3. Проблемы горных территорий и возможные способы их решения. Природные и антропогенные катаклизмы. Природные катаклизмы в горах представляют результат геотектонической природы гор и их экологических характеристик. Однако катаклизмы зачастую вызываются деятельностью человека. Перед ...
... обыкновенный, орлан-белохвост, сапсан, кречет, осетр сибирский, голец арктический, муксун, тугун, хариус сибирский. 3. Влияние динамики изменения площади особо охраняемых территорий на сохранение биоразнообразия 3.1 Динамика численности охотничье-промысловых животных В последние 30 лет все компоненты ландшафтов северной тайги Западной Сибири, включая орнитокомплексы, испытывают мощное ...
... , а целое – на части. Анализ и синтез. Антропный принцип. Принципы универсального эволюционизма. Вместе с представлением о коэволюции - составляющие эволюционно-синергетической парадигмы. Естествознание из ценностно-нейтрального знания превращается в аксиологически ориентированное, предполагающее введение этических, эстетических и т.п. норм в научное исследование. Биоэнергоинформатика – новое ...
0 комментариев