Навигация
Значение нуклеиновых кислот
69904
знака
2
таблицы
16
изображений
3.2. Значение нуклеиновых кислот
Значение нуклеиновых кислот очень велико. Особенности их химического строения обеспечивают возможность хранения, переноса в цитоплазму и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой клетке. Белки обусловливают большинство свойств и признаков клеток. Понятно поэтому, что стабильность структуры нуклеиновых кислот - важнейшее условие нормальной жизнедеятельности клеток и организма в целом. Любые изменения строения нуклеиновых кислот влекут за собой изменения структуры клеток или активности физиологических процессов в них, влияя таким образом на жизнеспособность.
Существует два типа нуклеиновых кислот: ДНК и РНК.
РНК (рибонуклеиновая кислота), так же как ДНК, представляет собой полимер мономерами которого служат нуклеотиды. Азотистые основания те же самые, что входят в состав ДНК (аденин, гуанин, цетозин); четвертое - урацил - присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дизоксирибозы другую пентозу - рибозу.
4. ДНК
4.1. Состав ДНК
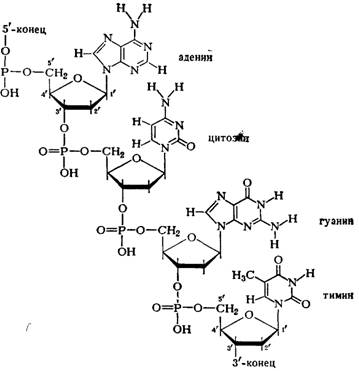
ДНК (дезоксирибонуклеиновая кислота) - биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин (А) или тимин (Т), цитозин (Ц) или гуанин (Г); пятиатомный сахар пентозу - дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов. В каждой цепи нуклеотиды соединяются путем образования ковалентных связей между дезоксирибозой одного и остатком фосфорной кислоты последующего нуклеотида. Объединяются две цепи в одну молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи.
Исследуя нуклеотидный состав ДНК различного происхождения, Чаргафф обнаружил следующие закономерности.
1. Все ДНК независимо от их происхождения содержат одинаковое число пуриновых и пиримидиновых оснований. Следовательно, в любой ДНК на каждый пуриновый нуклеотид приходится один пиримидиновый.
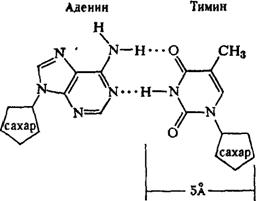
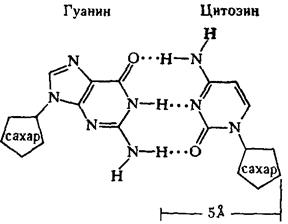
2. Любая ДНК всегда содержит в равных количествах попарно аденин и тимин, гуанин и цитозин, что обычно обозначают как А=Т и G=C. Из этих закономерностей вытекает третья.
3. Количество оснований, содержащих аминогруппы в положении 4 пиримидинового ядра и 6 пуринового (цитозин и аденин), равно количеству оснований, содержащих оксо-группу в тех же положениях (гуанин и тимин), т. е. A+C=G+T. Эти закономерности получили название правил Чаргаффа. Наряду с этим было установлено, что для каждого типа ДНК суммарное содержание гуанина и цитозина не равно суммарному содержанию аденина и тимина, т. е. что (G+C)/(A+T), как правило, отличается от единицы (может быть как больше, так и меньше ее). По этому признаку различают два основных типа ДНК: А Т-тип с преимущественным содержанием аденина и тимина и G C-тип с преимущественным содержанием гуанина и цитозина.
Величину отношения содержания суммы гуанина и цитозина к сумме содержания аденина и тимина, характеризующую нуклеотидный состав данного вида ДНК, принято называть коэффициентом специфичности. Каждая ДНК имеет характерный коэффициент специфичности, который может изменяться в пределах от 0,3 до 2,8. При подсчете коэффициента специфичности учитывается содержание минорных оснований, а также замены основных оснований их производными. Например, при подсчете коэффициента специфичности для ЭДНК зародышей пшеницы, в которой содержится 6% 5-метилцитозина, последний входит в сумму содержания гуанина (22,7%) и цитозина (16,8%). Смысл правил Чаргаффа для ДНК стал понятным после установления ее пространственной структуры.
4.2. Макромолекулярная структура ДНК
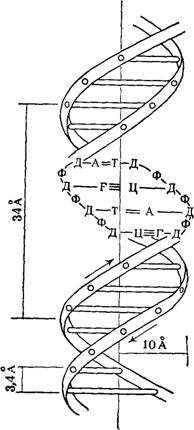
В 1953 г. Уотсон и Крик, опираясь на известные данные о конформаци нуклеозидных остатков, о характере межнуклеотидной связи в ДНК и закономерности нуклеотидного состава ДНК (правила Чаргаффа), расшифровали рентгенограммы паракристаллической формы ДНК [так называемой В-формы, образующейся при влажности выше 80% и при высокой концентрации противоионов (Li+) в образце]. Согласно их модели, молекула ДНК представляет собой правильную спираль, образованную двумя полидезоксирибонуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Диаметр спирали практически постоянен вдоль всей ее длины и равен 1,8 нм (18 А).
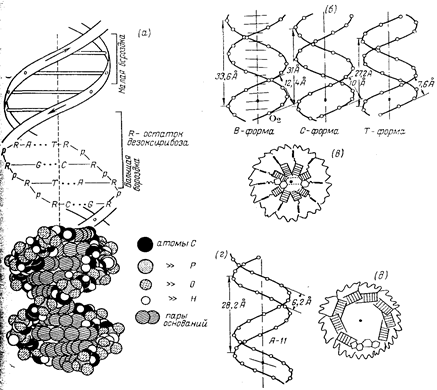
Макромолекулярная структура ДНК.
(а)—Модель Уотсона — Крика;
(6)—параметры спиралей В-, С- и Т-форм ДНК (проекции перпендикулярно оси спирали);
(в)—поперечный разрез спирали ДНК в В-форме (заштрихованные прямоугольники изображают пары оснований);
(г)—параметры спирали ДНК в А-форме;
(д)—поперечный разрез спирали ДНК в А-форме.
Длина витка спирали, который соответствует ее периоду идентичности, составляет 3,37 нм (33,7 А). На один виток спирали приходится 10 остатков оснований в одной цепи. Расстояние между плоскостями оснований равно, таким образом, примерно 0,34 нм (3,4 А). Плоскости остатков оснований перпендикулярны длинной оси спирали. Плоскости углеводных остатков несколько отклоняются от этой оси (первоначально Уотсон и .Крик предположили, что они параллельны ей).
Из рисунка видно, что углеводофосфатный остов молекулы обращен наружу. Спираль закручена таким образом, что на ее поверхности можно выделить две различные по размерам бороздки (их часто называют также желобками) — большую, шириной примерно 2,2 нм (22 А), и малую —шириной около 1,2 нм (12А). Спираль — правовращающая. Полидезоксирибонуклеотидные цепи в ней антипараллельны: это означает, что если мы будем двигаться вдоль длинной оси спирали от одного ее конца к другому, то в одной цепи мы будем проходить фосфодиэфирные связи в направлении 3'à5', а в другой — в направлении 5'à3'. Иными словами, на каждом из концов линейной молекулы ДНК расположены 5'-конец одной и 3'-конец другой цепи.
Регулярность спирали требует, чтобы против остатка пуринового основания в одной цепи находился остаток пиримидинового основания в другой цепи. Как уже подчеркивалось, это требование реализуется в виде принципа образования комплементарных пар оснований, т. е. остаткам аденина и гуанина в одной цепи соответствуют остатки тимина и цитозина в другой цепи (и наоборот).
Таким образом, последовательность нуклеотидов в одной цепи молекулы ДНК предопределяет нуклеотидную последовательность другой цепи.
Этот принцип является главным следствием модели Уотсона и Крика, поскольку он в удивительно простых химических терминах объясняет основное функциональное назначение ДНК — быть хранителем генетической информации.
Заканчивая рассмотрение модели Уотсона и Крика, остается добавить, что соседние пары остатков оснований в ДНК, находящейся в В-форме, повернуты друг относительно друга на 36° (угол между прямыми, соединяющими атомы С1' в соседних комплементарных парах).
Похожие работы
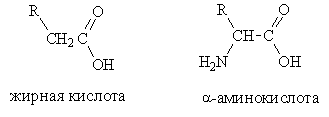


... частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка. Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и ...
... остатки, соединяющиеся водородными связями с комплементарными основаниями матричной цепи. Рост синтезирующейся цепи происходит в направлении 3'-ОН — 3'-ОН-концам, антипараллельно матричной цепи. Репликация ДНК приводит к удвоению количества генетического материала клетки и, как правило,— к клеточному делению. Поэтому репликация происходит тем чаще, чем короче время генерации вируса или бактерии и ...
... с агрессивным поведением иммунной системы. 21 Медицинская биотехнология, ее задачи и достижения. Биотехнология представляет собой область знаний, которая возникла и оформилась на стыке микробиологии, молекулярной биологии, генетической инженерии, химической технологии и ряда других наук. Рождение биотехнологии обусловлено потребностями общества в новых, более дешевых продуктах для ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
0 комментариев