Навигация
7.1. ДНК-полимеразы
В 1957 году А. Корнберг обнаружил у кишечной палочки фермент, катализирующий процесс полимеризации ДНК из нуклеотидов; он был назван ДНК-полимеразой. Затем ДНК-полимеразы выявили и в других организмах. Было показано, что субстратами всех этих ферментов служат дезоксирибонуклеозидтрифосфаты (дНТФ), полимеризующиеся на одноцепочной ДНК-матрице. ДНК-полимеразы последовательно наращивают одноцепочную цепь ДНК, шаг за шагом присоединяя к ней следующие звенья в направлении от 5-' к 3'-концу, причем выбор очередного дНТФ диктуется матрицей. Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом богатой энергией связи между первым и вторым фосфатными остатками в дНТФ и отщеплением пирофосфата, что делает реакцию в целом энергетически выгодной.
В клетках обычно присутствует несколько типов ДНК-полимераз, выполняющих различные функции и имеющих разное строение. Они могут быть построены из различного количества белковых цепей (субъединиц), от одной до десятков, однако все они работают на любых последовательностях нуклеотидов матрицы. Задача этих ферментов - сделать точную копию каждой матрицы.
7.2. Точность синтеза ДНК и механизм коррекции
Генетический материал живых организмов имеет огромные размеры и реплицируется с высокой точностью. В среднем в процессе воспроизведения генома млекопитающего, состоящего из ДНК длиной 3 миллиарда пар нуклеотидов, возникает не более трех ошибок. При этом ДНК синтезируется чрезвычайно быстро: скорость ее полимеризации колеблется в пределах от 500 нуклеотидов в секунду у бактерий, до 50 нуклеотидов в секунду у млекопитающих).
Высокая точность репликации, наряду с ее высокой скоростью, обеспечивается наличием специальных механизмов, осуществляющих коррекцию, то есть устраняющих ошибки. Суть механизма коррекции заключается в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний (3'-концевой) нуклеотид растущей цепи ДНК образовал правильную уотсон-криковскую пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии реакции произошло ошибочное спаривание оснований, то дальнейшая полимеризация останавливается до тех пор, пока ошибка не будет исправлена. Для этого фермент перемещается в обратном направлении и вырезает последнее добавленное звено, после чего его место может занять правильный нуклеотидпредшественник. Иными словами, многие (но не все) ДНК-полимеразы обладают, помимо 5'-3'-синтетической активности, еще и 3'-гидролизующей активностью, которая обеспечивает удаление ошибочно спаренных с матрицей нуклеотидов.
8. ОСНОВНЫЕ ПРИНЦИПЫ РЕПЛИКАЦИИ
Основные правила, в соответствии с которыми происходит репликация, были выяснены в опытах с бактериями, однако они справедливы также и для высших организмов.
8.1. Инициация цепей ДНК
ДНК-полимеразы не могут начинать синтеза ДНК на матрице, а способны только добавлять новые дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся полинуклеотидной цепи. Такую заранее образованную цепь, к которой добавляются нуклеотиды, называют затравкой. Короткую РНК- затравку синтезирует из рибонуклеозидтрифосфатов фермент, не обладающий корректирующей активностью и называемый ДНК-праймазой (от англ. primer - затравка). Праймазная активность может принадлежать либо отдельному ферменту, либо одной из субъединиц ДНК-полимеразы. Затравка, синтезированная этим неточным ферментом, не умеющим исправлять ошибки, отличается от остальной новосинтезированной цепи ДНК, поскольку состоит из рибонуклеотидов, и далее может быть удалена.
Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-полимеразой. Выполнившая свою функцию РНК-затравка удаляется специальным ферментом, а образованная при этом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3'-ОН-конец соседнего фрагмента. Удаление крайних РНК-праймеров, комплементарных 3'-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче на 10-20 нуклеотидов (у разных видов размер РНК-затравок различен). В этом заключается так называемая "проблема недорепликации концов линейных молекул". В случае репликации кольцевых бактериальных ДНК этой проблемы не существует, так как первые по времени образованиЯ РНК-затравки удаляются ферментом, который одновременно заполняет образующуюся брешь путем наращивания 3'-ОН-конца растущей цепи ДНК, направленной в "хвост" удаляемому праймеру. Проблема недорепликации 3'-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента - теломеразы. В 1985 году он был обнаружен у равноресничной инфузории Tetrahymena thermophila, а впоследствии - в дрожжах, растениях и животных, в том числе в яичниках человека.
Теломераза является ДНК-полимеразой, достраивающей 3'-концы линейных молекул ДНК хромосом короткими (6-8 нуклеотидов) повторяющимися последовательностями (у позвоночных TTAGGG). Согласно номенклатуре, этот фермент называют ДНК- нуклеотидилэкзотрансферазой или теломерной терминальной трансферазой. Помимо белковой части теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами. Длина теломеразной РНК колеблется от 150 нуклеотидов у простейших до 1400 нуклеотидов у дрожжей, у человека - 450 нуклеотидов. Сам факт наличия в молекуле РНК последовательности, по которой идет матричный синтез куска ДНК, позволяет отнести теломеразу к своеобразной обратной транскриптазе, то есть ферменту, способному вести синтез ДНК по матрице РНК.
В результате того что после каждой репликации дочерние цепи ДНК оказываются короче материнских на размер первого РНК-праймера (10-20 нуклеотидов), образуются выступающие однонитевые 3'-концы материнских цепей. Они-то и узнаются теломеразой, которая последовательно наращивает материнские цепи (у человека на сотни повторов), используя 3'-ОН-концы их в качестве затравок, а РНК, входящую в состав фермента, в качестве матрицы. Образующиеся длинные одноцепочные концы, в свою очередь, служат матрицами для синтеза дочерних цепей по традиционному репликативному механизму.
Постепенное укорочение ДНК хромосом во время репликации является одной из теорий "старения" клеточных колоний. Еще в 1971 году отечественный ученый А.М. Оловников в своей теории маргинотомии (от лат. marginalis -краевой, tome - сечение) предположил, что это явление лежит в основе ограниченного потенциала удвоения, наблюдаемого у нормальных соматических клеток. Американский ученый Леонард Хейфлик в начале 60-х годов показал, что если для культивирования взять клетки новорожденных детей, то они могут пройти 80-90 делений, в то время как соматические клетки от 70-летних делятся только 20- 30 раз. Ограничение на число клеточных делений и называют лимитом Хейфлика.
Похожие работы


... частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка. Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и ...
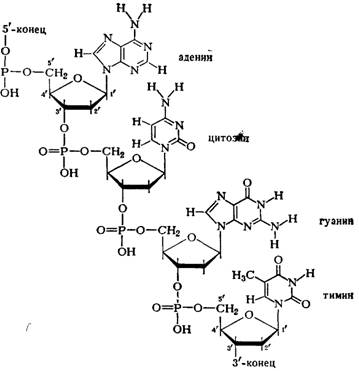
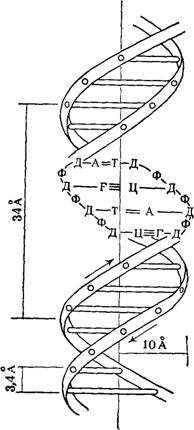
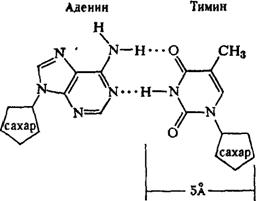
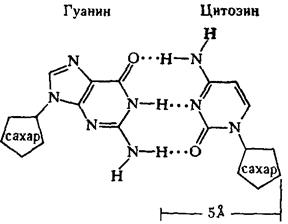
... остатки, соединяющиеся водородными связями с комплементарными основаниями матричной цепи. Рост синтезирующейся цепи происходит в направлении 3'-ОН — 3'-ОН-концам, антипараллельно матричной цепи. Репликация ДНК приводит к удвоению количества генетического материала клетки и, как правило,— к клеточному делению. Поэтому репликация происходит тем чаще, чем короче время генерации вируса или бактерии и ...
... с агрессивным поведением иммунной системы. 21 Медицинская биотехнология, ее задачи и достижения. Биотехнология представляет собой область знаний, которая возникла и оформилась на стыке микробиологии, молекулярной биологии, генетической инженерии, химической технологии и ряда других наук. Рождение биотехнологии обусловлено потребностями общества в новых, более дешевых продуктах для ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
0 комментариев