Протеиназы микробного происхождения
Ферменты, их физико-химические свойства
Расщепление коллагенсодержащего сырья и его применение
Определение величины рН
Идентификация протеиназ в ПААГ [16]
Фракционирование протеолитического комплексного
Исследование процессов кислотной и термической инактивации
Влияние ионов металлов и ингибиторов на активный центр фермента
Гидролиз коллагенсодержащего сырья: ноги птиц, шквара
Межпредметность – современный принцип обучения
Навигация
Исследование процессов кислотной и термической инактивации
Исследование некоторых физико-химических свойств протеиназы Penicillium wortmannii
99399
знаков
17
таблиц
8
изображений
3.5 Исследование процессов кислотной и термической инактивации.
Изучение термо- и рН-стабильности ферментов часто несёт прикладной характер.
Исследование этих характеристик проводится остаточной активностью фермента после выдержки его раствора в течение некоторого времени при определённых рН и температуре [11].
Нами были проведены исследования кинетики кислотной и термической инактивации протеиназ Penicillium wortmannii 2091 и рассчитаны кинетические параметры этого процесса.
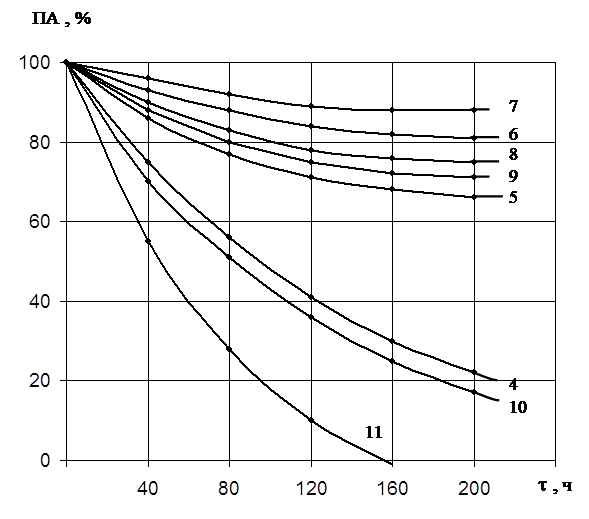
При изучении термо- и рН –стабильности раствор препарата выдерживали в фосфатном буфере в диапазоне рН от 5,0 до 12,0 и температур от 30 до 60оС. Периодически отбирали аликвотные доли раствора и определяли остаточную протеолитическую активность.
Полученные результаты по инактивации обоих ферментов показали, что протеиназа 1 сохраняет активность в широком диапазоне рН. При значении рН 7,0 через 200 часов фермент сохраняет около 90% активности (рис.4). При значениях рН 6,0 – 9,0 активность фермента снижается до 70 – 75%, это указывает на то, что фермент в указанном интервале не подвержен автолизу, а, следовательно, его нативная конформация обладает высокой стабильностью. При значениях рН ниже 6,0 и выше 11,0 каталитическая активность фермента быстро снижается.
Во всех случаях ПА/КлА-const, это свидетельствует о том, что мы имеем дело с одним ферментом.
Термическую инактивацию протеиназ изучали в интервале температур 30 – 60оС. Протеиназа 1 в области высокой рН-стабильности инактивируется почти полностью при температуре 60оС в течение 60 часов, в то время как протеиназа 2 при этой же температуре инактивируется полностью уже за 3 часа. Данные свидетельствуют о высокой термостабильности протеиназы 1. Инактивация протеиназы 2, происходящая при высокой температуре, по-видимому, определяется процессом разворачивания белковой глобулы.
Протеиназа 1, обладающая коллагенолитическим действием, нас интересует с практической точки зрения, поэтому были рассчитаны некоторые кинетические характеристики для этого фермента.
Если предположить, что в каждом элементарном акте процесса инактивации фермента под действием Н+ - ионов участвует одна его молекула, то кислотную инактивацию можно представить в виде реакции первого порядка. Кинетическое уравнение первого порядка имеет вид:
2,303 lg E0/E=K,
где Е0 – исходная активность фермента
Е – активность в момент времени
К – константа скорости инактивации, характеризующая потерю активности в течение часа, час-1.
 | |||
| |||
Остаточную активность выражали в процентах от исходной и затем использовали в расчётах констант инактивации. Величину находили, как среднее из 5 – 6 определений (табл.2).
Таблица 2.
Кислотная инактивация протеиназы 1 при температуре 500С.
| t, ч | Значения рН | |||||||
| 5,0 | 7,0 | 9,0 | 11,0 | |||||
| Е | К*103ч-1 | Е | К*103ч-1 | Е | К*103ч-1 | Е | К*103ч-1 | |
| 0 | 100 | 2 | 100 | 2 | 100 | 2 | 100 | 2 |
| 12 | 65,1 | 37,0 | 100 | 2 | 100 | 2 | 63,2 | 38,1 |
| 24 | 45,2 | 33,6 | 92,3 | 2,33 | 92,3 | 2,33 | 42,3 | 36,0 |
| 48 | 16,8 | 37,7 | 88,0 | 2,54 | 87,8 | 2,54 | 18,1 | 36,3 |
| 96 | 7,6 | 36,8 | 81,0 | 2,23 | 81,5 | 2,19 | 7,0 | 37,1 |
| 120 | 6,7 | 35,7 | 66,3 | 2,50 | 68,2 | 2,30 | 6,5 | 37,7 |
| 144 | 6,5 | 33,1 | 62,2 | 2,56 | 60,9 | 2,54 | 6,0 | 34,0 |
Как видно из табл.2, при определённом рН, значения констант достаточно близки друг к другу, максимальное отклонение от средних значений не превышает 10 – 15%, что вполне допустимо в исследованиях кинетики химических реакций. Это свидетельствует о том, что процесс инактивации протеиназы 1 является реакцией первого порядка. Различия в значениях при рН 5,0 и 11,0 и при рН 7,0 – 10,0 на целый порядок ещё раз указывают на лабильность фермента в слабо - кислой и слабо - щелочной зонах.
Исследования термической инактивации протеиназы 1 при различных значениях рН позволили рассчитать константы инактивации для температур 30, 40, 50, 60оС, а затем найти термодинамические параметры этого процесса.
Термодинамические расчёты были проведены только для вышеуказанных температур (табл.3).
Таблица 3.
Термодинамические характеристики активированного комплекса
протеиназы I.
| t, oC | PH | Еакт | DН¹ | DF¹ | DS Дж*К-1*моль-1 |
| Дж*моль-1 | |||||
| 30-60 | 5,0 | 245,8 | 244,6 | 53,2 | 600,3 |
| 30-40 | 7,0 | 75,6 | 72,0 | 68,1 | 13,6 |
| 40-60 | 7,0 | 300,5 | 298,2 | 60,0 | 734,9 |
| 30-40 | 9,0 | 72,4 | 71,0 | 69,9 | 9,9 |
| 40-60 | 9,0 | 295,9 | 294,1 | 60,0 | 723,9 |
| 30-40 | 11,0 | 255,9 | 253,6 | 54,9 | 624,3 |
Поскольку инактивация протеиназы была необратимой, для определения энтальпии DН¹, свободной энергии DF¹ и энтропии DS¹ воспользовались теорией абсолютных скоростей Эйринга.
При повышении температуры скорость инактивации возрастает. Это можно объяснить тем, что тепловая энергия разрушает гидрофобные взаимодействия, которые играют важную роль в стабильности белков. В результате происходит развёртывание полипептидной цепи, что подтверждается высокими значениями DS¹и согласуется с литературными данными.
Таким образом, изменение величины рН вызывает разрушение электростатических сил, и решающую роль в этих условиях в процессе инактивации играют, по-видимому, гидрофобные взаимодействия.
0 комментариев