Навигация
Синтез и расщепление гликогена
57980
знаков
0
таблиц
0
изображений
4. Синтез и расщепление гликогена
При повышении концентрации глюкозы в крови, например, в результате ее всасывания в кишечнике при пищеварении, увеличивается поступление глюкозы в клетки и по крайней мере часть этой глюкозы может быть использована для синтеза гликогена. Накопление резерва углеводов в клетках в виде гликогена имеет определенные преимущества перед накоплением глюкозы, так как не сопровождается повышением внутриклеточного осмотического давления. Вместе с тем, при недостатке глюкозы гликоген легко расщепляется до глюкозы или ее фосфорных эфиров, а образовавшиеся мономерные единицы используются клетками с энергетическими или пластическими целями.
4.1. Синтез гликогена
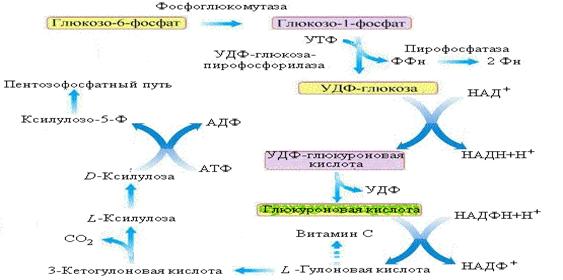
Поступившая в клетки глюкоза подвергается фосфорилированию при участии ферментов гексокиназы или глюкокиназы:
Затем гл-1-ф взаимодействует с уридинтрифосфатам с образованием УДФ-глюкозы при участии фермента УДФ-глюкозопирофосфорилазы [ или глюкозо-1-фосфатуридилтрансферазы ]:
Пирофосфат сразу расщепляется на два остатка фосфорной кислоты при участии фермента пирофосфатазы. Эта реакция сопровождается потерей энергии порядка 7 ккал/моль, в результате чего реакция образования УДФ-глюкозы становится необратимой - термодинамический контроль направления процесса.
На следующем этапе остаток глюкозы из УДФ-глюкозы переносится на синтезирующуюся молекулу гликогена при участии фермента гликогенсинтетазы: и молекула гликогена удлинняется на один остаток глюкозы. Фермент гликогенсинтетаза способна присоединить остаток глюкозы из УДФглюкозы к строящейся молекуле гликогена только путем образования a -1,4-гликозидной связи. Следовательно, при участии только одного этого фермента может быть синтезирован лишь линейный полимер. Гликоген же - полимер разветвленный и имеющиеся в молекуле разветвления формируются с участием другого фермента: амило- 1,4--> 1,6 - гликозилтрансферазы. Этот фермент, называемый иначе ферментом ветвления, переносит фрагмент из 5 - 7 мономерных звеньев с конца линейного участка синтезируемого полисахарида ближе к его средине, причем этот фрагмент присоединяется к полимерной цепи за счет образования a - 1,6-гликозидной связи:
Следует заметить, что по другим данным отщепляемый фрагмент, состоящий минимум из 6 глюкозных остатков, переносится на соседнюю цепочку строящегося разветвленного полисахарида. В любом случае в дальнейшем обе цепи удлинняются за счет действия гликогенсинтетазы, а новые разветвления формируются с участием фермента ветвления.
Синтез гликогена идет во всех органах и тканях, однако наибольшее содержание наблюдается в печени [ от 2 до 5-6% общей массы органа ] и в мышцах [ до 1 % от их массы ]. Включение 1 остатка глюкозы в молекулу гликогена сопровождается использованием 2 макроэргических эквивалентов ( 1 АТФ и 1 УТФ ), так что синтез гликогена в клетках может идти лишь при достаточной энергообеспеченности клеток.
4.2. Мобилизация гликогена
Гликоген, как резерв глюкозы, накапливается в клетках во время пищеварения и расходуется в постабсорбционном периоде. Расщепление гликогена в печени или его мобилизация осуществляется при участии фермента гликогенфосфоррилазы часто называемой просто фосфорилазой. Этот фермент катализирует фосфоролитическое расщепление a-1,4-гликозидных связей концевых остатков глюкозы полимера:
Для расщепления молекулы в районе разветвлений необходимы два дополнительных фермента: так называемый дебранчинг (деветвящий) фермент и амило-1,6-гликозидаза, причем в результате действия последнего фермента в клетках образуется свободная глюкоза, которая может или покинуть клетку, или подвергнуться фосфорилированию.
Гл-1-ф в клетках изомеризуется с участием фосфоглюкомутазы в гл-6-ф. Дальнейшая судьба гл-6-фосфата определяется наличием или отсутствием в клетках фермента глюкозо-6-фосфатазы. Если фермент присутствует в клетке, он катализирует гидролитическое отщепление от гл-6-фосфата остатка фосфорной кислоты с образованием свободной глюкозы: которая может проникать через наружную клеточную мембрану и поступать в кровяное русло. Если же глюкозо-6-фосфатазы в клетках нет, то дефосфорилирования глюкозы не происходит и глюкозный остаток может быть утилизирован только данной конкретной клеткой. Заметим, что расщепление гликогена до глюкозы не нуждается в дополнительном притоке энергии.
В большинстве органов и тканей человека глюкозо-6-фосфатаза отсутствует, поэтому запасенный в них гликоген используется лишь для собственных нужд. Типичным представителем таких тканей является мышечная ткань. Глюкозо-6-фосфатаза имеется лишь в печени, почках и кишечнике, но наиболее существенным является наличие фермента в печени ( точнее, в гепатоцитах ), т.к. этот орган выполняет роль своего рода буфера, поглощающего глюкозу при повышении ее содержания в крови и поставляющего глюкозу в кровь, когда концентрация глюкозы в крови начинает падать.
4.3. Регуляция процессов синтеза и распада гликогена
Сопоставив метаболические пути синтеза и мобилизации гликогена, мы увидим, что они различны:
Это обстоятельство дает возможность раздельно регулировать обсуждаемые процессы. Регуляция осуществляется на уровне двух ферментов: гликогенсинтетазы, участвующей в синтезе гликогена, и фосфорилазы, катализирующей расщепление гликогена.
Основным механизмом регуляции активности этих ферментов является их ковалентная модификация путем фосфорилирования-дефосфорилирования. Фосорилированная фосфорилаза или фосфорилаза "a" высокоактивна, в то же время фосфорилированная гликогенсинтетаза или синтетаза "b" неактивна. Таким образом, если оба фермента находятся в фосфорилированной форме, в клетке идет расщепление гликогена с образованием глюкозы. В дефосфорилированном состоянии, наоборот, неактивна фосфорилаза ( в форме "b") и активна гликогенсинтетаза ( в форме "a" ), в этой ситуации в клетке идет синтез гликогена из глюкозы.
Поскольку гликоген печени играет роль резерва глюкозы для всего организма, его синтез или распад должен контролироваться надклеточными регуляторными механизмами, работа которых должна быть направлена на поддержание постоянной концентрации глюкозы в крови. Эти механизмы должны обеспечивать включение синтеза гликогена в гепатоцитах при повышенных концентрациях глюкозы в крови и усиливать расщепление гликогена при падении содержания глюкозы в крови.
Итак, первичным сигналом, стимулирующим мобилизацию гликогена в печени, является снижение концентрации глюкозы в крови. В ответ на него альфа-клетки поджелудочной железы выбрасывают в кровь свой гормон - глюкагон. Глюкагон, циркулирующий в крови, взаимодействует со своим белком-рецептором, находящемся на внешней стороне наружной клеточной мембраны гепатоцита. образуя гор мон-рецепторный комплекс. Образование гормон-рецепторного комплекса приводит с помощью специального механизма к активации фермента аденилатциклазы, находящегося на внутренней поверхности наружной клеточной мембраны. Фермент катализирует образование в клетке циклической 3,5-АМФ ( цАМФ ) из АТФ.
В свою очередь, цАМФ активирует в клетке фермент цАМФ-зависимую протеинкиназу. Неактивная форма протеинкиназы представляет собой олигомер, состоящий из четырех субъединиц: 2 регуляторных и двух каталитических. При повышении концентрации цАМФ в клетке к каждой из регуляторных субъединиц протеинкиназы присоединяется по 2 молекулы цАМФ, конформация регуляторных субъединиц изменяется и олигомер распадается на регуляторные и каталитичес кие субъединицы. Свободные каталитические субъединицы катализирует фосфорилирование в клетке ряда ферментов, в том числе фосфорилирование гликогенсинтетазы с переводом ее в неактивное состояние, выключая таким образом синтез гликогена . Одновременно идет фос форилирование киназы фосфорилазы, а этот фермент, активируясь при его фосфорилировании, в свою очередь катализирует фосфорилирование фосфорилазы с переводом его в активную форму, т.е. в форму "a". В результате активации фосфорилазы включается расщепление гликогена и гепатоциты начинают поставлять глюкозу в кровь.
Попутно отметим, что при стимуляции расщепления гликогена в печени катехоламинами в качестве главных посредников выступают b - рецепторы гепатоцитов, связывающие адреналин. При этом происходит повышение содержания ионов Са в клетках, где они стимулируют Са/кальмодулинчувствительную киназу фосфорилазы, которая в свою очередь активирует фосфорилазу путем её фосфорилирования.
Повышение концентрации глюкозы в крови является внешним сигналом для гепатоцитов в отношении стимуляции синтеза гликогена и связывания таким образом излишней глюкозы из русла крови.
Срабатывает следующий механизм: при повышении концентрации глюкозы в крови возрастает и ее содержание в гепатоцитах. Повышение концентрации глюкозы в гепатоцитах, в свою очередь, достаточно сложным путем активирует в них фермент фосфопротеинфосфатазу, которая ка - тализирует отщепление от фосфорилированных белков остатков фосфорной кислоты. Дефосфорилирование активной фосфорилазы переводит ее в неактивную форму, а дефосфорилирование неактивной гликогенсинтетазы активирует фермент. В результате система переходит в состояние, обеспечивающие синтез гликогена из глюкозы.
В снижении фосфорилазной активности в гепатоцитах определенную роль играет гормон b-клеток поджелудочной железы инсулин. Он выделяется b-клетками в ответ на повышение содержания глюкозы в крови. Его связывание с инсулиновыми рецепторами на поверхности гепатоцитов приводит к активации в клетках печени фермента фосфодиэстеразы, катализирующего превращение цАМФ в обычную АМФ, не обладающую способность стимулировать образование активной протеинкиназы. Этим путем прекращается нарабатывание в гепатоцитах активной фосфорилазы, что также имеет значение для ингибирования расщепления гликогена.
Вполне естественно, что механизмы регуляции синтеза и распада гликогена в клетках различных органов имеют свои особенности. В качестве примера можно указать, что в миоцитах покоящихся мышц или мышц, выполняющих небольшую по интенсивности работу, практически нет фосфорилазы "a", но расщепление гликогена все же идет. Дело в том, что мышечная фосфорилаза, находящаяся в дефосфорилированном состоянии или в форме "b", является аллостерическим ферментом и активируется имеющимися в миоцитах АМФ и неорганическим фосфатом. Активированная таким образом фосфорилаза "b" обеспечивает скорость мобилизации гликогена, достаточную для выполнения умеренной физической работы.
Однако при выполнении интенсивной работы, в особенности если нагрузка резко возрастает, этого уровня мобилизации гликогена становится недостаточно. В таком случае срабатывают надклеточные механизмы регуляции. В ответ на внезапно возникшую потребность в интенсивной мышечной деятельности в кровь поступает гормон адреналин из мозгового вещества надпочечников. Адреналин, связываясь с рецепторами на поверхности мышечных клеток, вызывает ответную реакцию миоцитов, близкую по своему механизму к только что описанной реакции гепатоцитов на глюкагон. В мышечных клетках появляется фосфорилаза "a" и инактивируется гликогенсинтетаза, а образовавшийся гл-6-ф используется как энергетическое "топливо", окислительный распад которого обеспечивает энергией мышечное сокращение.
Следует заметить, что высокие концентрации адреналина, наблюдающиеся в крови людей в условиях эмоционального стресса, ускоряют расщепление гликогена в печени, повышая тем самым содержание глюкозы в крови - защитная реакция, направленная на экстренную мобилизация энергетических ресурсов.
Похожие работы
... механизмах синтеза гликогена, во всех окислительных путях превращения глюкозы и в синтезе других моносахаридов,необходимых для клетки. Место, которое занимает данная реакции в обмене глюкозы позволяет ее счиатать ключевой реакцией обмена углеводов. Гексокиназная реакция необратима (G= -16,7 кДж/моль), поэтому для превращения глюкозо-6-фосфата в свободную глюкозу в клетках печени и почек ...

... α,d – глюкоза глюкозо – 6 – фосфат С образованием глюкозо – 6 – фосфата пути гликолиза и гликогенолиза совпадают. Глюкозо – 6 – фосфат занимает ключевое место в обмене углеводов. Он вступает в следующие метаболические пути: глюкозо – 6 – фосфат глюкоза + Н3РО4 фруктозо – 6 – фосфат пентозный путь распада (поступает в кровь и др. ...
... . Динамика химических превращений, происходящих в клетках, изучается биологической химией. Задачей физиологии является определение общих затрат веществ и энергии организмом и того, как они должны восполняться с помощью полноценного питания. Энергетический обмен служит показателем общего состояния и физиологической активности организма. Единица измерения энергии, обычно применяемая в биологии и ...
... затратах энергии; 2) при переменных затратах энергии и 3) при затратах на синтез продукции. Наибольшее количество теплоты образуется в органах с интенсивным обменом веществ и большой массой – печени и мышцах. При мышечной работе химическая энергия только на треть переходит в механическую работу, остальные две трети переходят в теплоту. Теплопродукция может увеличиваться в 3…5 раз за счет ...
0 комментариев