Обзор литературы
Материал и методы исследования
Методы исследования
Генетико – статистические методы анализа
Распространенность полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN среди здоровых лиц (контрольная группа)
Анализ связи полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN с туберкулезом
Анализ накопления случаев туберкулеза в семьях больных
Анализ связи полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN с патогенетически важными параметрами болезни
Анализ ассоциаций исследованных генов с количественными признаками туберкулеза
Навигация
Анализ связи полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN с туберкулезом
Аллельные варианты генов-кандидатов подверженности туберкулезу у русского населения Западной Сибири
191682
знака
23
таблицы
7
изображений
3.2 Анализ связи полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN с туберкулезом
Туберкулез – инфекционное заболевание, обладающее клиническим полиморфизмом. Специфическая гранулема – субстрат болезни – может развиться в любом органе или ткани [Cтруков А.И., Кауфман О.Я., 1989; Ерохин В.В., Земскова З.С., 2003]. Современная классификация туберкулеза, в основу которой положено два признака: локализация процесса и клинико – рентгенологические особенности форм, насчитывает 23 основных клинических формы туберкулеза [Хоменко А.Г., 1996]. Такое разнообразие проявлений одного заболевания определяют множество факторов. Несомненно, в этом случае играют немаловажную роль свойства самой микобактерии туберкулеза, такие как вирулентность, резистентность к противотуберкулезным препаратам, а также массивность инфекции. От состояния естественных защитных сил организма (барьерная функция слизистой оболочки дыхательных путей и желудочно-кишечного тракта) также будет зависеть исход встречи человека и микобактерии туберкулеза.
Результаты современных генетических исследований не оставили сомнений в существовании наследственной предрасположенности к туберкулезу [Хоменко А.Г., 1996; Чуканова В.П., 2001; Hill A.V.S., 1999]. Они показали, что одной из причин, определяющих такое разнообразие клинических проявлений, является полиморфизм генов, чьи продукты экспрессии вовлечены в патогенез заболевания. В то же время, пока не ясно, какие именно варианты генов имеют решающее значение. В связи с этим важно изучить вклад конкретных сочетаний аллелей в развитие болезни.
В исследованной выборке 304 больных туберкулезом русских жителей г. Томска и Томской области проанализировано распределение генотипов и частот аллелей полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN (табл. 10). Выявлено отклонение наблюдаемого распределения генотипов полиморфизма B/b гена VDR от ожидаемого при РХВ (χ2=6,95; р=0,008) за счет повышенной гетерозиготности (табл. 10). Учитывая неслучайный характер отбора индивидов в выборке, можно предположить наличие вероятного вклада гена VDR в патогенез туберкулеза. Для исследованных полиморфизмов генов NRAMP1, IL12B, IL1B, IL1RN в общей группе больных туберкулезом отклонение от РХВ не показано.
Известно, что у жителей Тувы больных туберкулезом выявлено отклонение от РХВ для полиморфизмов 1465-85G/A и D543N гена NRAMP1 за счет пониженного количества гетерозигот в обоих случаях и для полиморфизмов B/b и F/f гена VDR за счет повышенной гетерозиготности [Рудко А.А., 2004]. Таким образом, как для русских, так и для тувинцев, больных туберкулезом, характерно повышенное количество гетерозигот B/b гена VDR. Сравнение частот аллелей и генотипов у русских и тувинцев, больных туберкулезом, показало статистически значимые отличия для всех исследованных полиморфных маркеров (табл. 11). Максимальная степень отличий частот аллелей и генотипов показана для полиморфизма B/b гена VDR.
Таблица 10 Частоты аллелей и генотипов исследованных полиморфизмов у больных туберкулезом
| Ген | Поли-морфизм | Гено-типы | N.O. | N.E. | Частота аллеля | χ2 (d.f.) | Hobs | Hexp | D |
| NRAMP1 | 469+14 G/C | GG GC CC | 179 94 6 | 183,07 85,86 10,07 | G= 0,810 | 2,25 (1) | 0,337 | 0,308 | +0,095 |
| D543N | DD DN NN | 263 14 1 | 262,23 15,54 0,23 | D= 0,971 | 1,28 (1) | 0,050 | 0,056 | -0,099 | |
| 1465-85 G/A | GG GA AA | 126 126 27 | 128,03 121,94 29,03 | G= 0,677 | 0,25 (1) | 0,452 | 0,437 | +0,033 | |
| 274C/T | CC CT TT | 163 122 14 | 167,82 112,37 18,81 | C= 0,749 | 2,01 (1) | 0,408 | 0,376 | +0,086 | |
| IL12B | 1188 A/C | AA AC CC | 162 100 17 | 161,09 101,82 16,09 | A= 0,760 | 0,05 (1) | 0,358 | 0,365 | -0,018 |
| VDR | B/b | BB Bb bb | 49 168 76 | 60,37 145,26 87,37 | b= 0,546 | 6,95* (1) | 0,573 | 0,496 | +0,157 |
| F/f | FF Ff ff | 124 134 40 | 122,42 137,16 38,42 | F= 0,641 | 0,12 (1) | 0,450 | 0,460 | -0,023 | |
| IL1B | +3953 A1/A2 | A1A1 A1A2 A2A2 | 169 108 24 | 165,21 115,57 20,22 | A1 0,741 | 1,16 (1) | 0,359 | 0,384 | -0,066 |
| IL1RN | VNTR | A1A1 A1A2 A1A3 A1A4 A2A2 A2A3 другие | 158 100 10 1 25 4 1 | 152,43 110,16 11,53 0,43 19,90 4,17 0,38 | A1= 0,714 A2= 0,258 A3= 0,027 | 6,35 (6) | 0,385 | 0,422 | -0,088 |
Примечание. Обозначения см. табл. 6
Таблица 11 Сравнение частот аллелей и генотипов у русских и тувинцев с ТБ
| Полимор-физм | Гено-типы | Частоты (%) | p | Ал-лели | Частоты | p | ||
| русские | тувинцы | русские | тувинцы | |||||
| NRAMP1 | ||||||||
| 469+14 G/C | GG GC CC | 179 (64,2) 94 (33,7) 6 (2,1) | 198 (85) 32 (13,7) 3 (1,3) | 0,000 | G | 0,810 | 0,918 | 0,000 |
| D543N | DD DN NN | 263 (94,6) 14 (5) 1 (0,4) | 191 (80,9) 39 (16,5) 6 (2,6) | 0,000 | D | 0,971 | 0,892 | 0,000 |
| 1465-85G/A | GG GA AA | 126 (45,2) 126 (45,2) 27 (9,6) | 153 (65,7) 63 (27) 17 (7,3) | 0,000 | G | 0,677 | 0,794 | 0,000 |
| 274C/T | CC CT TT | 163 (54,5) 122 (40,8) 14 (4,7) | 190 (80,5) 45 (19,1) 1 (0,4) | 0,000 | C | 0,749 | 0,900 | 0,000 |
| IL12B | ||||||||
| 1188 A/C | AA AC CC | 162 (58) 100 (35,8) 17 (6,2) | 90 (38,6) 102 (43,8) 41 (17,6) | 0,000 | A | 0,760 | 0,605 | 0,000 |
| VDR | ||||||||
| B/b | BB Bb bb | 49 (16,7) 168 (57,3) 76 (26) | 3 (1,3) 82 (35) 149 (63,7) | 0,000 | b | 0,546 | 0,812 | 0,000 |
| F/f | FF Ff ff | 124 (41,6) 134 (45) 40 (13,4) | 125 (53,6) 102 (43,8) 6 (2,6) | 0,000 | F | 0,641 | 0,755 | 0,000 |
| IL1B | ||||||||
| +3953 A1/A2 | A1A1 A1A2 A2A2 | 169 (56,1) 108 (35,9) 24 (8) | 185 (79,1) 47 (20) 2 (0,9) | 0,000 | A1 | 0,741 | 0,891 | 0,000 |
Примечание. р – достигнутый уровень значимости при сравнении частот аллелей и генотипов
У русских с ТБ и здоровых по сравнению с жителями Тувы чаще наблюдали гомозиготный по аллелею В и гетерозиготный генотипы. Среди жителей г. Томска, больных туберкулезом, по сравнению с тувинцами статистически значимо чаще встретились аллели 469+14C, 1465-85A, 274T гена NRAMP1, f гена VDR, и +3953А2 гена IL1В, реже регистрировались аллели 543N гена NRAMP1 и 1188С гена IL12B. В целом можно отметить, что отличия в распределении генотипов и аллелей между русскими и тувинцами были идентичными у больных и здоровых: у русских с туберкулезом преобладали те же аллели генов, что и у здоровых.
При сравнении распределений генотипов и частот аллелей в выборках больных и здоровых лиц, живущих в г. Томске и Томской области, выявлены статистически значимые отличия для полиморфизма 274С/Т гена NRAMP1 и трех генов, принимающих участие в продукции интерлейкинов (табл. 12). Так, у больных туберкулезом статистически значимо чаще регистрировался гетерозиготный генотип 274С/Т гена NRAMP1 (OR=1,66, 95%CI:1,02–2,71; p=0,040). Лица, обладающие гомозиготным генотипом 274С/C чаще встречались среди здоровых (OR=0,54, 95%CI:0,33–0,87; р=0,010). Показаны отличия частот аллелей у больных и здоровых для полиморфизма +3953 А1/А2 гена IL1B (p=0,036) за счет повышения доли аллеля А2 у больных туберкулезом. В тоже время, не найдено отличий при сравнении распределения генотипов этого полиморфизма гена IL1В (табл. 12).
Различия по VNTR полиморфизму гена IL1RN в выборках больных и здоровых касались как частот аллелей, так и частот генотипов. Аллель А2 статистически значимо чаще встречался у пациентов с туберкулезом. При сравнении частот генотипов выявлено более высокое количество гетерозигот А1/А2 у больных туберкулезом по сравнению с контролем (OR=2,10, 95%CI:1,26–3,51, р=0,003) (табл. 12). В противоположность этому у африканцев гетерозиготы по аллелю А2 IL1RN статистически значимо реже встречались у больных туберкулезом, чем в контрольной группе [Bellamy R., Ruwende C., 1998].
Найдена ассоциация полиморфизма 1188А/С гена IL12В с туберкулезом. Установлено, что распределение генотипов и частота аллелей в группах больных и здоровых различаются. В выборке больных статистически значимо чаще встречаются гомозиготы по аллелю 1188С (р=0,035). В данном случае вероятность заболеть туберкулезом обладателей гомозиготного генотипа 1188С/С значимо выше, чем у индивидов с гомозиготным генотипом 1188А/А и гетерозиготным (OR=8,31, 95%CI:1,15–169,30, р=0,030) (табл. 12). Несмотря на отклонение распределения генотипов от ожидаемого при РХВ полиморфизма B/b гена VDR у больных туберкулезом, не показано отличий между больными и здоровыми.
Диагноз туберкулеза устанавливается на основании наличия у больного характерных жалоб, эпидемиологического анамнеза, анамнеза заболевания, клинических симптомов, характерных изменений в общем анализе крови и рентгенологической картины. Однако только при обнаружении микобактерий туберкулеза (МБТ) в мокроте диагноз туберкулеза можно считать верифицированным [Рабухин А.Е.,1976].
Учитывая это, при дальнейшем анализе из общей группы больных была выделена выборка пациентов с бактериовыделением (МБТ+) (n=234). При сравнении распределения генотипов и частот аллелей в группах больных лабораторно подтвержденным туберкулезом и контрольной, найдены статистически значимые отличия для полиморфизма 274С/Т гена NRAMP1, аллельного варианта +3953 А1/А2 гена IL1B, VNTR полиморфизма гена IL1RN и полиморфизма 1188А/С IL12В (табл. 13). Частота аллеля 274Т гена NRAMP1 была выше в группе больных (р=0,009). Показана протективная роль генотипа 274С/С (OR=0,53, 95%CI: 0,32– 0,88, р=0,012).
Таблица 12 Статистические показатели для сравнения частот аллелей и генотипов у больных туберкулезом и здоровых
| Ген | Полимор-физм | Гено-типы | Сравнение | OR (95%CI) | |
| Генотипов р | Аллелей р | ||||
| NRAMP1 | 469+14 G/C | GG GC CC | 0,393 | 0,229 | 0,74 (0,46–1,18) 1,32 (0,83–2,13) 1,48 (0,27–10,77) |
| D543N | DD DN | 0,283 | 0,377 | 1,66 (0,70–3,88) 0,56 (0,24–1,34) | |
| 1465-85 G/A | GG GA AA | 0,134 | 0,312 | 0,70 (0,45–1,08) 1,54 (0,99–2,41) 0,86 (0,42–1,76) | |
| 274C/T | CC CT TT | 0,009 | 0,020 | 0,54* (0,33–0,87) 1,66* (1,02–2,71) 2,80 (0,59–18,13) | |
| IL12B | 1188 A/C | AA AC CC | 0,035 | 0,044 | 0,72 (0,45–1,13) 1,12 (0,70–1,78) 8,31* (1,15–169,30) |
| VDR | B/b | BB Bb bb | 0,925 | 0,791 | 0,94 (0,51–1,76) 0,96 (0,60–1,54) 1,10 (0,64–1,91) |
| F/f | FF Ff ff | 0,705 | 0,469 | 1,20 (0,75–1,93) 0,89 (0,57–1,41) 0,88 (0,46–1,69) | |
| IL1B | +3953 A1/A2 | A1A1 A1A2 A2A2 | 0,108 | 0,036 | 0,70 (0,45–1,08) 1,21 (0,77–1,90) 2,32 (0,82–7,10) |
| IL1RN | VNTR | A1A1 A1A2 A2A2 | 0,030 | 0,023 | 0,57* (0,36–0,88) 2,10* (1,26–3,51) 0,97 (0,45–2,13) |
Примечание. р - достигнутый уровень значимости; OR – величина отношения шансов; (95%CI) - доверительный интервал для OR; * - р < 0,05
Таблица 13 Статистические показатели для сравнения частот аллелей и генотипов у больных лабораторно подтвержденным туберкулезом и здоровых
| Полимор-физм | Гено-тип | N | р | OR (95% СI) | Частота аллеля | р |
| NRAMP1 | ||||||
| 274С/Т | CC CT TT | 126 94 12 | 0,021 | 0,53* (0,32–0,88) 1,64* (0,99–2,73) 3,11 (0,64–20,47) | C=0,746 | 0,009 |
| IL12B | ||||||
| 1188 А/С | AA AC CC | 125 82 12 | 0,044 | 0,69 (0,43–1,11) 1,20 (0,74–1,94) 7,42* (0,99–154,58) | A=0,758 | 0,046 |
| IL1B | ||||||
| +3953 А1/А2 | A1A1 A1A2 A2A2 | 131 82 21 | 0,080 | 0,69 (0,44–1,09) 1,16 (0,73–1,87) 2,64 (0,91–8,21) | A1=0,735 | 0,036 |
| IL1RN | ||||||
| VNTR | A1A1 A1A2 A2A2 другие | 121 79 21 11 | 0,042 | 0,55* (0,35–0,87) 2,16* (1,28–3,68) 1,06 (0,48–2,38) - | А1=0,707 A2=0,267 A3=0,024 A4=0,002 | 0,029 |
Примечание. р - достигнутый уровень значимости; OR – величина отношения шансов; (95%CI) - доверительный интервал для OR; N – количество индивидов с определенным генотипом; *- p < 0,05
Рисковыми для туберкулеза оказались генотипы 274С/Т и 274Т/Т гена NRAMP1. Не найдено отличий в распределении частот генотипов +3953 А1/А2 полиморфизма гена IL1B, в то время как аллель А2 статистически значимо чаще встречался в группе больных лабораторно подтвержденным туберкулезом, чем в группе здоровых индивидов (р=0,036) (табл. 13). Ассоциация VNTR полиморфизма гена IL1RN, а имменно связь А2 аллеля с туберкулезом, прослеживается как в общей выборке больных, так и в группе пациентов с наличием МБТ в мокроте. При анализе результатов подсчета отношения шансов показана более высокая подверженность туберкулезу индивидов с генотипом А1/А2 (OR=2,16, 95%CI:1,28–3,68, p=0,003). Протективным эффектом обладал полиморфизм А1А1 гена IL1RN (OR=0,55).
Для полиморфизма 1188А/С гена IL12B выявлены различия между больными туберкулезом МБТ+ и здоровыми по частотам аллелей (р=0,046) и генотипов (р=0,044). У больных туберкулезом чаще, чем в контрольной группе регистрировался гомозиготный генотип 1188С/С, риск развития туберкулезного процесса у этих индивидов был в семь раз выше, чем у обладателей других генотипов этого полиморфизма.
Различают первичный туберкулез, который развивается в ранее не инфицированном МБТ организме, а также вторичный генез туберкулеза, когда заболевание возникает при повторном инфицировании. Микобактерии туберкулеза проникают в организм различными путями. При небольшом их количестве, слабой вирулентности и при достаточной устойчивости организма они могут не вызвать специфических изменений на месте внедрения, а также и при лимфо–гематогенном рассеивании. Возникает состояние латентного микробизма [Земскова З.С., Дорожкова И.Р., 1984]. В дальнейшем может развиться инфицирование МБТ, когда возбудитель находится в макроорганизме, но человек не заболевает туберкулезом. При других условиях в результате первичного заражения образуются туберкулезные очаги в том или ином органе (первичный туберкулез).
Если же на фоне инфицирования в организм дополнительно попадет новая порция вирулентных микобактерий, может развиться заболевание, которое будет иметь вторичный генез (вторичный туберкулез). Наличие факторов, приводящих к снижению общей и местной резистентности организма, таких как курение, сахарный диабет, кортикостероидная терапия, лечение цитостатиками, ВИЧ-инфицирование и другие, увеличит вероятность этого события. Также вторичный туберкулез может возникнуть в результате обострения перенесенного в прошлом первичного процесса.
Патогенетические механизмы первичных и вторичных форм туберкулеза разнятся. Поэтому различаются и клинические формы, характерные для этих двух групп. Если для первичного туберкулеза свойственно поражение лимфатических узлов средостения или мезентериальных и, как осложнение, легочной ткани (исключение составляет первичный туберкулезный комплекс, при котором первичный аффект формируется в легком), то при вторичном генезе туберкулеза, как правило, поражается легочная ткань.
В связи с этим фактом можно предположить, что резистентность макроорганизма к первичному и вторичному туберкулезу обусловлена разными факторами иммунитета, а значит, варианты генов, приводящие к предрасположенности к разным по генезу формам туберкулеза, также будут различаться. В ходе дальнейшего анализа больные туберкулезом были поделены на две группы в зависимости от того, на каком этапе симбиоза человека и микобактерии произошло заболевание. В выборку больных с первичным туберкулезом были отнесены пациенты, у которых выставили диагнозы: туберкулез внутригрудных лимфоузлов (n=36), первичный туберкулезный комплекс (n=3), гематогенно-диссеминированный туберкулез легких первичного периода (n=2), плеврит туберкулезной этиологии первичного периода (n=2). Сравнение частот аллелей и распределений генотипов в выборках больных первичным туберкулезом и контрольной показало наличие ассоциации с заболеванием полиморфизма 1465-85G/A гена NRAMP1: найдены статистически значимые различия частот аллелей (р=0,047) и генотипов (р=0,004) этого полиморфизма в группах больных и здоровых. У заболевших первичным туберкулезом чаще, чем в контрольной группе, встречался гетерозиготный генотип 1465-85G/A, риск заболевания у этих индивидов был три раза выше, чем у обладателей других генотипов этого полиморфизма (OR=3,16, 95%CI:1,47–6,86; р=0,002) (табл. 14). Показано, что генотип 1465-85G/G чаще регистрировался у здоровых индивидов (OR=0,33, 95%CI:0,15–0,73; р=0,005), таким образом, установлена его протективная роль в отношении первичного туберкулеза.
Таблица 14 Cтатистические показатели для сравнения частот аллелей и генотипов полиморфизма 1465-85 G/A гена NRAMP1 у больных туберкулезом первичного периода и здоровых
| Полиморфизм | Гено-тип | N | р | OR (95%CI) | Частота аллеля | р |
| 1465-85 G/A | GG GA AA | 12 27 4 | 0,004 | 0,33* (0,15–0,73) 3,16* (1,47–6,86) 0,82 (0,22–2,86) | G=0,593 | 0,047 |
Примечание. Обозначения см. табл. 13
Известно, что продукт гена Nramp1 у мышей участвует в процессе фагоцитоза, влияя на судьбу МБТ в макроорганизме. Кроме того, на мышах показано, что именно ранняя фаза инфекции с микобактериями туберкулеза штамма BCG находится под контролем гена Nramp1 [Scamene E. et al., 1980]. По всей видимости, дефект продукции или функции человеческого гомолога Nramp1 при первой встрече человека с микобактерией может способствовать возникновению заболевания. Тогда, как в организме инфицированного ранее индивида срабатывают механизмы иммунологической защиты, и мутации в гене, а следовательно и дефекты белка NRAMP1 не оказывают подобного влияния.
Ранее показано, что полиморфизм 1465-85G/A гена NRAMP1 оказывает предрасполагающую роль к инфильтративному туберкулезу у тувинцев [Рудко А.А., 2004]. Частота гомозигот А/А этого полиморфизма была практически в два раза выше у больных ТБ, чем у здоровых. Учитывая результаты исследования генетических основ подверженности к туберкулезу русских и тувинцев, можно отметить, что ген NRAMP1 оказывает влияние на предрасположенность к заболеванию, проявляя этническую специфичность.
Выборку больных с вторичным происхождением туберкулеза составили пациенты с установленным диагнозом инфильтративного туберкулеза легких (150 человек), диссеминированного туберкулеза легких (65 человек), очагового туберкулеза (27 человек), казеозной пневмонии (5 человек), впервые выявленного фиброзно–кавернозного туберкулеза легких (4 человека), туберкуломы (4 человека), туберкулеза почечной паренхимы (3 человека), туберкулеза бронха (2 человека), плеврита туберкулезной этиологии (1 человек). Всем больным диагноз туберкулеза был установлен впервые.
При сравнении частот аллелей и распределений генотипов ассоциацию с вторичным туберкулезом продемонстрировали полиморфизм 274С/Т гена NRAMP1, полиморфный маркер гена IL1В, VNTR полиморфизм гена IL1RN, а также ген IL12B (табл. 15). Аллель 274Т гена NRAMP1 чаще встречался в группе больных (р=0,005), причем у лиц гетерозиготных по этому полиморфизму риск заболеть туберкулезом возрастал в 1,75 раза (95%СI:1,06–2,88; p=0,026), а у гомозигот 274Т/Т шанс заболеть был в пять раз выше, чем у обладателей других генотипов этого полиморфизма (95%CI:1,07–33,72; p=0,037).
Показано, что генотип 274С/С статистически значимо чаще встречался у здоровых лиц (OR=0,51, 95%CI:0,31–0,82; p=0,005). В целом, сравнивая результаты, полученные при исследовании частот аллелей и генотипов у больных первичным и вторичным туберкулезом, можно отметить, что разные полиморфизмы гена NRAMP1 оказывают неравнозначное влияние на предрасположенность к туберкулезу у русских г. Томска.
Таблица 15 Статистические показатели для сравнения частот аллелей и генотипов у больных вторичным туберкулезом легких и здоровых
| Полимор-физм | Гено-тип | N | р | OR (95% СI) | Частота аллеля | р |
| NRAMP1 | ||||||
| 274С/Т | CC CT TT | 136 108 13 | 0,010 | 0,51* (0,31–0,82) 1,75* (1,06–2,88) 5,01* (1,07–33,72) | C=0,739 | 0,005 |
| IL1B | ||||||
| +3953 А1/А2 | A1A1 A1A2 A2A2 | 146 92 21 | 0,103 | 0,70 (0,45–1,10) 1,19 (0,75–1,89) 2,36 (0,82–7,34) | A1=0,741 | 0,041 |
| IL1RN | ||||||
| VNTR | A1A1 A1A2 A2A2 другие | 134 89 23 12 | 0,025 | 0,55* (0,35–0,86) 2,20* (1,31–3,72) 1,04 (0,48–2,31) - | А1=0,705 A2=0,269 A3=0,023 A4=0,003 | 0,019 |
Примечание. р - достигнутый уровень значимости; OR – величина отношения шансов; (95%CI) - доверительный интервал для OR; N – количество индивидов с определенным генотипом; *- p < 0,05
Показано, что полиморфизм 1465-85 G/A гена NRAMP1 проявил связь с первичным туберкулезом. Учитывая, что при аналогичном сравнении частот генотипов и аллелей в выборках больных вторичными формами туберкулеза и здоровых, различий не было найдено, полученные данные позволяют предположить наличие влияния этого полиморфизма на более раннее возникновение заболевания. Полиморфный вариант 274С/Т оказался ассоциированным с вторичным генезом заболевания. Для маркеров D543N и 469+14G/C гена NRAMP1 не показано предрасполагающего влияния на ТБ у русских жителей г. Томска.
Известно, что IL–1β является провоспалительным цитокином, а значит, его варианты могут приводить к ослаблению защитной реакции организма и, как следствие, способствовать заболеванию при наличии возбудителя в организме. Выявлены различия частот аллелей при сравнении группы контроля и больных вторичным туберкулезом, при которых заболевание ассоциированно с аллелем А2 IL1В (χ2=4,17, р=0,041). Продукт гена IL1RN является рецепторным антагонистом IL–1β, выполняя функцию специфического блокатора биологического действия IL–1β. Показано, что аллель А2 приводит к увеличению продукции белка IL–1RN [Wilkinson R. J. et al., 1999].
Выявлено статистически значимое увеличение частоты аллеля А2 VNTR полиморфизма гена IL1RN у больных по сравнению с контролем (χ2=9,92, р=0,019). Распределение генотипов этого полиморфизма также различалось в группах лиц с вторичным туберкулезом и здоровых (χ2=14,49, р=0,025): у больных чаще регистрировался гетерозиготный генотип А1/А2, риск развития заболевания у них был выше более чем в 2 раза, по сравнению с лицами с другими генотипами этого полиморфизма (95%CI: 1,31–3,72, р=0,002). Протективная роль в развитии вторичного туберкулеза показана для генотипа А1/А1, так как он статистически значимо чаще встречен у здоровых (OR=0,55, 95%CI: 0,35–0,86, р=0,007).
В целом, сравнение общей группы больных туберкулезом, выборки пациентов–бактериовыделителей, больных вторичным ТБ с контрольной выборкой показало аналогичные результаты, то есть связь полиморфизмов 274С/Т гена NRAMP1, +3953 А1/А2 гена IL1B и VNTR гена IL1RN с заболеванием. Общую выборку больных на 86% составили пациенты с вторичным туберкулезом. При первичном туберкулезе, как правило, не встречается выделение МБТ с мокротой [Рабухин А.Е., 1976]. В исследовании жителей г. Томска только один больной с первичным туберкулезным комплексом был бактериовыделителем. Так, выборка больных туберкулезом МБТ+ практически на 100% состояла из пациентов с туберкулезом вторичного периода. Таким образом, полиморфизм 274С/Т гена NRAMP1, +3953 А1/А2 полиморфизм гена IL1B и VNTR полиморфизм IL1RN проявили ассоциацию с вторичным туберкулезом. Тем более, что ассоциации, найденные при сравнении первичного туберкулеза с контролем, затрагивали иные гены.
Найдена связь полиморфизма 1188А/С гена IL12B как с первичным, так и с вторичным туберкулезом, причем генотип 1188С/С чаще встречался у больных, чем у здоровых (табл. 16). Это наблюдение вполне объяснимо, поскольку известно, что IL-12 участвует в активации макрофагов посредством Т-хелперов, вырабатывающих INF-γ, а активизированные макрофаги, в свою очередь, устремляются к месту нахождения микобактерий и активно их поглощают. Вероятно, варианты гена IL12B влияют на подверженность заболеванию у неинфицированных МБТ и инфицированных ранее людей.
Кроме анализа связи генетических маркеров с заболеванием по типу случай–контроль был использован метод оценки ассоциации болезни с генетическими маркерами на семейном материале с помощью Transmission Disequilibrium Test (TDT). Метод оценивает предпочтительность переноса аллеля М1 больному потомку от М1М2 гетерозиготных родителей.
Если аллель М1 переносится существенно больше, чем в половину раз, можно заключить, что маркерный локус сцеплен с локусом предрасположенности к заболеванию, и что аллель М1 положительно ассоциирован с аллелем, который увеличивает восприимчивость к заболеванию, или является таковым [Spielman R.S. et. al., 1993].
Таблица 16 Статистические показатели для сравнения частот аллелей и генотипов полиморфизма 1188А/С гена IL12B у больных различным по генезу туберкулезом и здоровых
| Выборка больных ТБ | Генотипы 1188А/С | Сравнение | OR(95%CI) | p | ||
| генотипов | аллелей | |||||
| ТБ первичного периода | АА | 27 | р=0,012 | р=0,370 | 0,93(0,42–2,06) | 0,997 |
| АС | 11 | 0,71(0,30–1,64) | 0,500 | |||
| СС | 4 | 13,47(1,35–326,42) | 0,013 | |||
| ТБ вторичного периода | АА | 135 | р=0,041 | р=0,041 | 0,69(0,43–1,10) | 0,120 |
| АС | 89 | 1,20(0,75–1,94) | 0,491 | |||
| СС | 13 | 7,43(1,00–153,87) | 0,023 | |||
Примечание. Обозначения см. табл. 12
Расчет TDT провели для всех исследованных полиморфных маркеров, за исключением D543N гена NRAMP1, для которого TDT не был рссчитан из-за низкой гетерозиготности родителей пробандов (табл. 17). Анализ показал наличие связи полиморфизма 1188А/С гена IL12B с туберкулезом. Дети, больные туберкулезом, лишь в 13% случаев наследовали от родителей аллель 1188А, тогда как аллель 1188С – в 87%.
Таблица 17 Число аллелей, унаследованных больными потомками от гетерозиготных родителей
| Ген | Полиморфизм | Аллели | Количество перенесенных аллелей | TDT | p (d.f.=1) |
| NRAMP1 | 469+14G/C | G | 10 | 1,00 | 0,317 |
| С | 6 | ||||
| 1465-85G/A | G | 12 | 0,15 | 0,695 | |
| A | 14 | ||||
| 274C/T | C | 9 | 0,06 | 0,808 | |
| T | 8 | ||||
| IL12B | 1188A/C | A | 2 | 8,07 | 0,005 |
| C | 13 | ||||
| VDR | B/b | B | 6 | 2,00 | 0,157 |
| b | 12 | ||||
| F/f | F | 11 | 0,00 | 1,000 | |
| f | 11 | ||||
| IL1B | +3953A1/A2 | A1 | 9 | 0,05 | 0,818 |
| A2 | 10 | ||||
| IL1RN | VNTR | A1 | 9 | 0,60 | 0,439 |
| не А1* | 6 |
Примечание. р – достигнутый уровень значимости; d.f. – число степеней свободы; * - аллели А2 и А3
Найденная связь изменчивости гена IL12B закономерна, поскольку продукт экспрессии этого гена – IL–12 является ключевым цитокином для усиления клеточно-опосредованного иммунного ответа и инициации эффективной противоинфекционной защиты. Он секретируется макрофагами в ответ на индукцию микробными компонентами и продуктами, активирует дифференцировку Т-лимфоцитов, повышает их цитотоксическую активность, усиливает пролиферацию естественных киллеров, Т-лимфоцитов и продукцию других цитокинов. Главный эффект – индукция синтеза INF-γ. Характер течения и исход многих инфекций зависят от способности возбудителя индуцировать синтез IL-12 [Тотолян А.А., Фрейдлин И.С., 2000]. Таким образом, у русских жителей г. Томска установлена ассоциация полиморфизма 1188А/С гена IL12B с заболеванием: индивиды-носители аллеля 1188С в большей степени подвержены туберкулезу, как первичному, так и вторичному. Важным обстоятельством с точки зрения достоверности является тот факт, что показанная связь маркера с заболеванием выявлена как методом случай – контороль, так и TDT на семейном материале.
Похожие работы
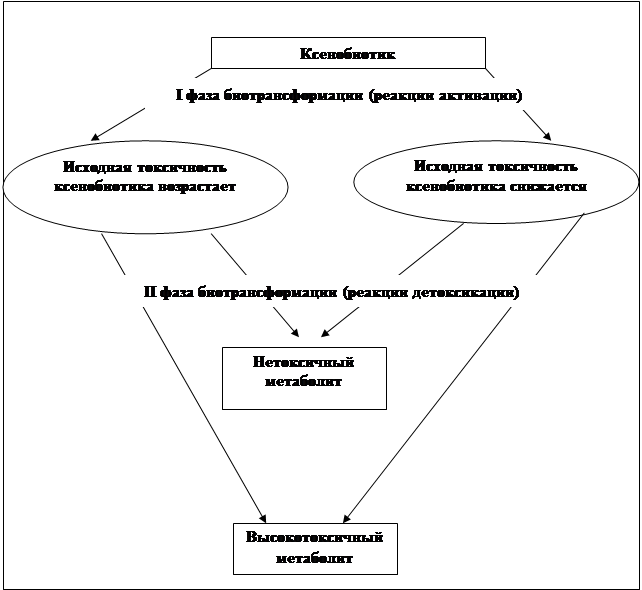
... препаратов. Установлена связь полиморфизма 313A>G гена GSTP1 с изменчивостью уровня аланинаминотрансферазы (р=0,021). 7. Выявлены различия в структуре генетической подверженности к бронхиальной астме и туберкулезу по генам ферментативной системы метаболизма ксенобиотиков: гены GSTM1, CYP2E1 и CYP2C19 связаны с бронхиальной астмой и значимыми для заболевания качественными и ...
0 комментариев