Теоретичні основи генно-модифікованих продуктів
ГЕНЕТИЧНИЙ КОД
СТРУКТУРНА ОРГАНІЗАЦІЯ ГЕНОМА
ГЕННО-ІНЖЕНЕРНІ ПІДХОДИ ДО СТВОРЕННЯ ІНТЕНСИВНИХ ТЕХНОЛОГІЙ В ХАРЧОВІЙ ГАЛУЗІ
ВИКОРИСТАННЯ ГЕННОЇ ІНЖЕНЕРІЇ В М’ЯСНІЙ ТА МОЛОЧНІЙ ПРОМИСЛОВОСТІ
ВИКОРИСТАННЯ ГЕННОЇ ІНЖЕНЕРІЇ У РОСЛИННИЦТВІ
МЕТОДИ ГЕННОЇ ІНЖЕНЕРІЇ МІКРООРГАНІЗМІВ
ПОЗИТИВНІ ТА НЕГАТИВНІ НАСЛІДКИ ВИКОРИСТАННЯ ГМП
МЕТОДИ ЗАСТОСУВАННЯ ГЕНЕТИЧНОЇ ІНЖЕНЕРІЇ В МЕДИЦИНІ
ГЕНЕТИЧНА ІНЖЕНЕРІЯ В ЕПІДЕМІОЛОГІЇ
ПРОБЛЕМИ ВИКОРИСТАННЯ ГЕНЕТИЧНО МОДИФІКОВАНОЇ СИРОВИНИ
Навигация
ГЕНЕТИЧНИЙ КОД
Теоретичні основи генно-модифікованих продуктів
131587
знаков
1
таблица
5
изображений
1.2 ГЕНЕТИЧНИЙ КОД
З моделі структури ДНК, запропонованої Уотсоном і Кріком, відомо, що генетична інформація передається за допомогою якоїсь специфічної послідовності нуклеотидів її молекули. Вперше питання про код було поставлене Гамовим у 1953 р.
Початок прямого генетико-біохімічного аналізу кодонів було покладено в 1961 р. Ніренбергом та Маттеї, які створили найпростіші синтетичні полімери й замінили ними нативні молекули іРНК в системі компонентів клітин бактерій та суміші амінокислот. У суміші кожного типу одна з амінокислот була помічена радіоактивним вуглецем С14, інші дев'ятнадцять не мали позначення. Було встановлено, що синтетичний полірибонуклеотид, складений тільки з урацилу (поліуридилова кислота - УУУУУ), визначає синтез білка, в якому кожна амінокислоти була фенілаланіном. Виходячи з цього, встановили, що триплет УУУ є кодоном для фенілаланіну.
У наступних подібних експериментах було визначено, що поліаденінова кислота містить код для лізину, а поліцитидинова - для проліну. Наступні досліди, де як штучні ІРНК виступали полінуклеотиди змішаного складу (наприклад, полі-АУ), виявили зв'язок між рядом інших поєднань нуклеотидів та конкретними амінокислотами.
Синтетичні полінуклеотиди створюють з використанням ферменту полінуклеотидфосфорилази, який зв'язує нуклеотиди у випадковому порядку. Для перших експериментів цього було достатньо, оскільки в них використовували синтетичні полінуклеотиди, складені з одного типу нуклеотидів. Потім були знайдені шляхи складніших синтезів молекул з різних нуклеотидів із різними положеннями.
Нову методику широко використав Очоа із співробітниками, що дало їм можливість визначити триплети для всіх 20 амінокислот (табл.4). При цьому було зазначено, що код має вироджений характер, який означає здатність для однієї й тієї ж кислоти бути кодованою кількома різними триплетами. Наприклад, є амінокислоти, які мають по шість кодонів; п'ять амінокислот, кожна з яких кодується чотирма різними кодонами. Так, аланін кодується триплетами ГЦУ, ГЦЦ, ГЦА, ГЦГ, куди в усіх випадках входять нуклеотиди цитозину та гуаніну. Поряд з тим є амінокислоти, що кодуються трьома, двома і тільки дві - одним триплетом азотистих основ.
Крім того, триплети УАА, УАГ, УГА не кодують амінокислоти, а є своєрідними "крапками" в процесі зчитування інформації. Якщо процес синтезу наближається до такої "крапки" в молекулі ДНК, синтез даного поліпептидного ланцюга припиняється. Після "крапки" починає синтезуватися нова молекула білка. Процес зчитування інформації відбувається в одному і тому ж напрямі. Так, якщо в молекулах азотисті основи розміщені в такому порядку: ААА, ЦЦЦ, УГУ, УЦУ, то це значить, що закодовані такі послідовно розміщені амінокислоти: лізин (ААА), пролін (ЦЦЦ), цистин (УГУ), серін (УЦУ). Саме в цій послідовності вони повинні розміщуватися в поліпептидному ланцюзі при синтезі білка. Якщо в першому триплеті ІРНК буде втрачено один аденін, то порядок основ набуває вигляду ААЦЦЦУГУУЦУ. Внаслідок цього склад всіх триплетів змінюється. Так, перший стане не ААА, а ААЦ. Подібний триплет кодує амінокислоту аспарагін, а не лізин, як було раніше. Другий стане вже не ЦЦЦ, а ЦЦГ і так далі. У деяких умовах іn vitro код може бути двозначним, тобто один триплет може кодувати кілька амінокислот. Кодон УУУ в звичайних умовах кодує амінокислоту фенілаланін. Проте якщо рибосоми обробити стрептоміцином, то цей кодон починає також кодувати ізолейцин і серин. Знижена температура та висока концентрація іонів Мg++ також зумовлюють двозначність у дії кодонів.
Таблиця
Генетичний код послідовності нуклеотидів у триплетних кодонах інформаційної РНК, який визначає певну амінокислоту
| Перший | Третій нуклеотид | (на 3'- кінці | кодону) | |||||
| нуклеотид (на 5'-кінці кодону) | Другий гуклео-тид | У | ц | А | г | |||
| У | У | Фен | Фен | Лей | Лей | |||
| ц | Сер | Сер | Сер | Сер | ||||
| А | Тир | Тир | Термінатор (знак закінчення) | Термінатор (знак закінчення) | ||||
| Г | Цис | Цис | Арг | Три | ||||
| У | Лей | Лей | Лей | Лей | ||||
| Ц | Ц | Про | Про | Про | Про | |||
| А | Гіс | Пс | Глі | Глі | ||||
| Г | Арг | Арг | Арг | Арг | ||||
| У | Іле | Іле | Іле | Мет | ||||
| А | Ц | Тре | Тре | Тре | Тре | |||
| А | Асп | Асп | Ліз | Ліз | ||||
| Г | Сер | Сер | Арг | Арг | ||||
| У | Вал | Вал | Вал | Вал | ||||
| г | Ц | Ала | Ала | Ала | Ала | |||
| А | Асп | Асп | Глу | Глу | ||||
| Г | Глі | Глі | Глі | Глі | ||||
Генетичний код характеризується неперекритістю. Цей принцип був доведений дослідженням мутацій, які порушують синтез білків. У випадку перекриття коду зміна в одній парі нуклеотидів неминуче повинна спричинити порушення в трансляції трьох амінокислот, бо у коді, що перехрещується, кожний з нуклеотидів входить до трьох кодонів. Насправді експериментами доведено, що мутації змінюють транслювання тільки однієї амінокислоти, що чітко вказує на неперекритість коду. Для поняття принципів генетичного коду значний інтерес являють дані по заміщенню амінокислот внаслідок зміни всередині триплетів.
У Е. соlі (кишкової палички, яка є в кишечнику всіх організмів) були одержані шість мутантів, що мали різні дефекти у триптофансинтетазі, причиною яких стали заміщення гліцину в специфічній точці поліпептидного ланцюга. Ці мутації відрізняються від попереднього стану зміною одного нуклеотиду. Якщо ця гіпотеза правильна, то вона може бути перевірена в дослідах по рекомбінації, оскільки кросинговер всередині триплетів повинен змінювати його код. Наприклад, схрещування між мутацією В, кодуючою аргінін, та мутацією С, кодуючою валін, повинні дати рекомбінантні триплети, які будуть кодувати серин та гліцин: мутант В - УГЦ -аргінін; мутант С - УУГ - гліцин.
У випадку кросинговеру на іншій ділянці (ГЦ і УГ) повинні з'явитися рекомбінанти - перший блок кросинговеру: ЦГГ - дикий тип - гліцин; другий -додатковий кросинговер: УУЦ - мутант - серин. У наступних дослідах ці рекомбінанти були виявлені і повністю відповідали передбаченням.
У випадках серповидноклітинності гемоглобіну у людини серед 300 амінокислот, які входять до складу гемоглобіну, тільки одна замінена мутацією. У гемоглобіні С в цьому місці з'являється інша амінокислота - лізин (рис.1)
(Численними дослідженнями встановлена дивовижна універсальність генетичного коду. Він однаково проявляє себе в системах, одержаних з вірусів, бактерій, водоростей та ссавців. Очевидно, він єдиний для всього органічного світу, що є одним з найпереконливіших доказів загального походження всієї живої природи.
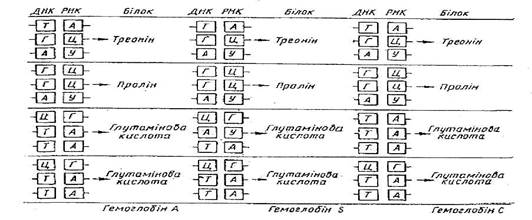
Рис. 1 Зміна нуклеотидів у кодових триплетах у цистроні, що визначає молекулу гемоглобіну у людини
Похожие работы
... . Більш як 40 країн, у тому числі й Україна, підписали Картагенський протокол. Не підписали його лише США, як і Конвенцію про біобезпеку. Треба зазначити, що реакція на продукти з генетично модифікованих джерел їжі різна в США та Європі. Споживачі в США виражають в основному позитивне ставлення до генної інженерії; Під час національного соціологічного опитування, проведеного Міжнародною радою з ...
... до цього, однак різниця в антигенах іноді становиться причиною серйозних захворювань, які несуть загрозу нормальному розвитку вагітності і життю дитини. Частіше всього зустрічається серологічний конфлікт по резус-фактору, який може привести до небезпечних порушень, особливо до гемолітичної хвороби плоду і новонародженого. Кожна людина з моменту свого зачаття має генетично визначену групу крові. І ...


... ЄТЬСЯ, що Одкровення було записано близько 66 року н.е. і, імовірно, доповнене Іоанном згодом через 30 років. З тих пір не проходило жодного століття (а в наш час і жодного року) без нових досліджень і тлумачень цього пророцтва. Число разючих збігів із пророкуванням Іоанна в кожнім столітті було велике, іноді навіть доходило до критичної маси, коли віруючі тієї чи інша країни готувалися до "кінця ...
... дає змогу суттєво розширити виробничі можливості працівників, збільшуючи плідність їх зусиль. У сучасних умовах вона є могутнім фактором розвитку продуктивної сили праці. 4.НАУКОВО-ТЕХНІЧНИЙ ПРОГРЕС ЯК ОСНОВА РОЗВИТКУ ВИРОБНИЦТВА І ЗНИЖЕННЯ ЙОГО ПОТЕНЦІАЛЬНОЇ НЕБЕЗПЕКИ 4.1.Екологічне становище України. Загострення проблем безпеки навколишнього середовнща і населення стало ...
0 комментариев