Теоретичні основи генно-модифікованих продуктів
ГЕНЕТИЧНИЙ КОД
СТРУКТУРНА ОРГАНІЗАЦІЯ ГЕНОМА
ГЕННО-ІНЖЕНЕРНІ ПІДХОДИ ДО СТВОРЕННЯ ІНТЕНСИВНИХ ТЕХНОЛОГІЙ В ХАРЧОВІЙ ГАЛУЗІ
ВИКОРИСТАННЯ ГЕННОЇ ІНЖЕНЕРІЇ В М’ЯСНІЙ ТА МОЛОЧНІЙ ПРОМИСЛОВОСТІ
ВИКОРИСТАННЯ ГЕННОЇ ІНЖЕНЕРІЇ У РОСЛИННИЦТВІ
МЕТОДИ ГЕННОЇ ІНЖЕНЕРІЇ МІКРООРГАНІЗМІВ
ПОЗИТИВНІ ТА НЕГАТИВНІ НАСЛІДКИ ВИКОРИСТАННЯ ГМП
МЕТОДИ ЗАСТОСУВАННЯ ГЕНЕТИЧНОЇ ІНЖЕНЕРІЇ В МЕДИЦИНІ
ГЕНЕТИЧНА ІНЖЕНЕРІЯ В ЕПІДЕМІОЛОГІЇ
ПРОБЛЕМИ ВИКОРИСТАННЯ ГЕНЕТИЧНО МОДИФІКОВАНОЇ СИРОВИНИ
Навигация
СТРУКТУРНА ОРГАНІЗАЦІЯ ГЕНОМА
Теоретичні основи генно-модифікованих продуктів
131587
знаков
1
таблица
5
изображений
1.3 СТРУКТУРНА ОРГАНІЗАЦІЯ ГЕНОМА
Під геномом розуміють сукупність носіїв спадкової інформації, що міститься в клітині. ДНК прокаріот оточена слабо зв’язаним з нею основним білком і ніяк структурно не організована. Характерна морфологія еукаріотичних хромосом свідчить, що вони організовані значно складніше, ніж: геноми прокаріотичних клітин. У еукаріотів більша частина ДНК знаходиться у міцному комплексі з білками та утворює нуклеопротеїдні волокна, які називаються хроматином. У переважній більшості клітин хроматинові білки представлені гістонів. Гістоні –цс невеликі за розміром (50-200 амінокислотних залишків) основні білки з позитивним зарядом (зумовлений наявністю трьох амінокислот: аргініну, лізину, гістидину). Утворення комплексу з ДНК (що має негативний заряд) відбувається за рахунок іонних зв’язків між; фосфатною групою полінуклеотидного ланцюга та аміногрупою поліпептиду. Гістоні розділяють на п’ять типів: Н1, Н2А, Н2В, НЗ, Н4, які відрізняються один від одного кількістю амінокислот та відношенням лізин : аргінін. Цікаво, що структура гістонів НЗ та Н4 з проростків гороху і з тімусу теляти, як довели Сміт та Де Ланж, дуже подібні, тобто послідовність амінокислот збереглася протягом приблизно 3-6*108 років з часу розділення всього живого на рослини та тварини. Ця консервативність свідчить, що зазначені гістони виконують дуже важливу функцію, яка виникла на початку еволюції еукаріот і збереглася до нашого часу. Білкам властива здатність до зміни заряду, форми молекул, до утворення водневих зв’язків, що може мати важливе значення у регуляції доступності ДНК до реплікації та транскрипції.
Слід зазначити, що, на відміну від ядерної ДНК, ДНК мітохондрій та хлоропластів не зв’язана з гістонів (як ДНК прокаріот), що підтверджує гіпотезу симбіотичного походження цих органоїдів.
Виходячи з даних, одержаних різними методами, Корнберг (1974) висловив припущення, що хроматин складається з повторюваних субодиниць, кожна з яких містить 200 пар нуклеотидів та по дві молекули гістонів Н2А, Н2В, НЗ, Н4. Повторювані одиниці називають нуклеосомами. Більша частина ДНК намотана на гістонову серцевину, решта, так звана міжнуклеосомна ДНК, з’єднує сусідні нуклеосоми та забезпечує гнучкість гістонів нитки. Таким чином, хроматинова нитка являє собою гнучкий ланцюг гістонів, що нагадує намистини на нитці.
В 60-х роках цього століття почались інтенсивні дослідження по локалізації конкретних генів на хромосомах для побудови хромосомних карт. Як встановлено, в геномі людини нараховується близько 500 тис. різних структурних генів, тобто генів, в яких закодована інформація про амінокислотну послідовність білків; кількість ДНК в ядрі клітини така, що відповідає числу генів, яка в 50-100 раз більша.
Значна кількість ДНК приходиться на повторювані послідовності нулеотидів, які можуть грати в хромосомі регуляторну, структурну або функціональну роль.
Реєструючи порушення, які викликаються мутаціями генів, вдалося виявити більше 900 генних локусів із загального числа 500 тис. структурних генів (геном людини), до якого входять гени, які кодують всі ферменти внутрішнього метаболізму, всі структурні білки та всі білки, яким властиві спеціальні функції, такі як гемоглобіни та імуноглобуліни, а також ферменти, які беруть участь в утворенні фібрил колагену (її роколагенпептидаза, глюкозилтрансфераза, лізнлоксидаза) та в утворенні нуклеїнових кислот (ДНК- та РНК-полімераза).
Істотна частина постульованої кількості генів приходиться на регуляторні гени.
Положення генних локусів на хромосомах людини визначали на основі аналізу гетерозигот та вивчення родин (варіації в межах виду), а пізніше – за допомогою гібридних клітин, які утворюються в результаті злиття соматичних клітин. Зручним інструментом досліджень був також метод гібридизації нуклеїнових кислот: таким засобом на хромосомах 13, 14, 15, 21 та 22 були локалізовані гени 183 та 283 рРНК, а на хромосомі 1 –гени 53 рРНК. Цим же методом встановлено, що гени а- та р-ланцюгів гемоглобіну знаходяться на хромосомах 2 та 4 або 5. Метод гібридизації нуклеїнових кислот дозволяє картувати гени гістонів, імуноглобулінів та колагенів.
Клонування нуклеїнових кислот та наступна гібридизація їх з фрагментами хромосом також можуть бути корисними для точної локалізації генів та побудови докладної карти геному.
Результати подібних досліджень будуть досить важливими для прогресу медицини, який буде залежати від повного розуміння функціонування геному. Дійсно, ряд хвороб пов’язані з наявністю дефектів певних генів. Лікування або попередження генетичних порушень потребує загальних знань структури, функціонування та регуляції роботи генів.
1.4 КЛОНУВАННЯ ГЕНІВ
Основою проведення генно-інженерних досліджень є молекула ДНК, що показана на схемах (рис. 2-3 ).При цьому роботи виконують в певній послідовності : спочатку виділяють гени з окремих клітин або синтезують їх поза організмом, потім включають нові гени у вектор, поєднують ДНК гена і вектора і одержують рекомбінантну ДНК; далі переносять визначені гени в геном клітини-хазяїна, проводять копіювання і розмноження виділених або синтезованих генів у складі вектора (клонування генів) і одержують генний продукт шляхом експериментальної експресії чужорідного гена в реципієнтній клітині. Відомо два шляхи виділення генів та створення ргкомбінантної ДНК.
Перший - за допомогою хімічного синтезу, а другий, більш поширений, грунтується на використанні особливих ферментів (рестриктаз), які мають властивість розпізнавати чужорідну ДНК, що проникла в організм, і розщеплювати її у відповідних ділянках. В результаті утворюються фрагменти різноманітних розмірів, які різняться між собою за довжиною. Відомо близько 500 ферментів рестриктаз і кожний розщеплює ДНК специфічно. Хоча багато з них за специфічністю подібні, проте кількість сайтів (ділянок) розщеплення становить близько 120. Зазначені ферменти позбавлені видової специфічності. Завдяки цьому можна поєднувати в одне ціле фрагменти ДНК будь-якого походження і подолати природний видовий бар'єр. На рисунку 5 схематично зображено дію ферменту на молекулу ДНК, що зумовлює відокремлення від неї частини нуклеотидів.
Частини й розриви ниток ДНК лігизують (склеюють) за допомогою ферменту лігази. Особливістю виділених ділянок нуклеотидів (генів) є так звані липкі кінці, через що їх можна приєднати до ділянок ДНК плазмід (для рослин і бактерій) або фагів (тварин). Таким чином створюється вектор для перенесення виділених генів у клітину-реципієнт.
Відомо інший шлях одержання фрагментів ДНК з липкими кінцями. Для цього виділені або штучно синтезовані ділянки ДНК обробляють ендонуклеазою, яка укорочує її з обох боків. Потім за допомогою ферменту полінуклеотидтрансферази добудовують до цих кінців ділянки аденінових і тимідинових нуклеотидів. Одержану молекулу рекомбінантної ДНК використовують для перенесення чужорідного гена в бактеріальну клітину. Така схема була використана для генів інсуліну, інтерферону, імуноглобуліну.
Молекули ДНК, які мають власний апарат реплікації і здатні доставляти в клітину потрібні гени, реплікувати їх, були названі векторами. Найбільш поширені вектори – це різноманітні плазміди, які часто спостерігаються у бактерій. Вектори для клітин ссавців будуються на основі вірусів, адено- та ретровірусів.
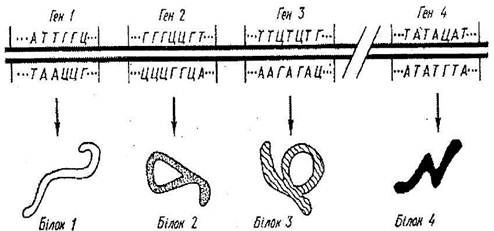
Рис.3 Молекула ДНК, поділена на гени
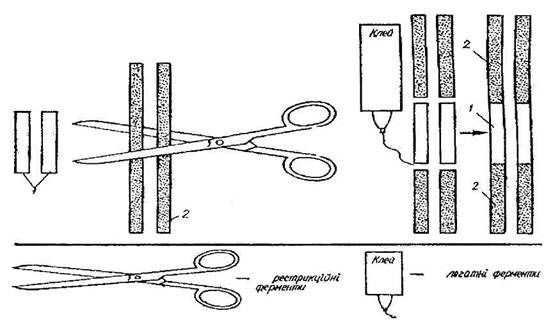
Рис.4 Процес утворення рекомбінантної ДНК
1-бактеріальна ДНК; 2-рослинна ДНК.
Потрібно враховувати, що наявність навіть введення гена у хромосому організма-хазяїна ще не дає можливості одержувати продукти його синтезу.
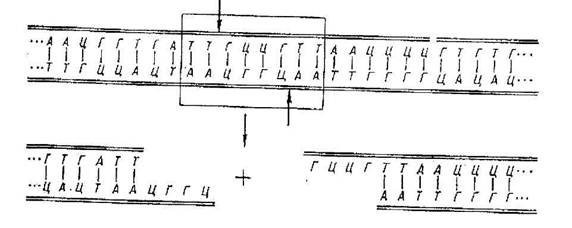
Рис.5 Рестрикційні ферменти розпізнають послідовності, виділені прямокутником, і вирізують ДНК на місці, зазначеному стрілками
Для того, щоб ген міг функціонувати, він повинен поряд з частиною, де закодована інформація, мати ще регуляторну ділянку. Це, так звані промотор та термінатор. З промотора починається зчитування інформації (транскрипція), а в термінаторі закодовано закінченння транскрипції з даного гена. Нині створено цілий «арсенал» клонованих промоторів, які дають можливість забезпечити проявлення генів у різних типах клітині.
Слід враховувати також, що не всі молекули плазмідної ДНК можуть мати вставки чужої ДНК і відповідно не будуть рекомбінантними. Більшість плазмід відновлює вихідну кільцеву структуру. Тому, перш за все необхідно відібрати бактерії, що містять рекомбінантні плазміди.
Для відбору рекомбінантних ДНК найбільш поширеною є система, при якій чужорідну ДНК вбудовують в частину плазмідного гена, що кодує стійкість проти певного антибіотика, наприклад, ампіциліну. У випадку вбудовування чужорідної ДНК цей ген перестає нормально функціонувати, що свідчить про наявність рекомбінантної ДНК.
і Молекули рекомбінантної плазміди розмножуються в клітині. В процесі ділення бактеріальної клітини вони розподіляються між дочірними клітинами і в кожній з них знову відновлюють свою кількість. В результаті створюються колонії бактерій, кожна з яких містить багато копій рекомбінантної ДНК. У кожному такому клоні міститься лише один відрізок ДНК тварини або рослини, який випадково потрапив у вихідну бактерію.
При цьому такий клон містить 1-2 гени, а якщо врахувати, що клонів значна кількість, то вони теоретично представляють всі гени, що є в геномі тварини.
Отже, для створення банку генів кроля, що характеризує всю молекулу ДНК або весь геном, необхідно 920000 клонів, для банку класичного об'єкта генетичної інженерії - кишкової палички - 1300. Для генома ссавців потрібен банк генів з фрагментами ДНК 0,8-1 млн. клонів)
Перший банк генів було створено для Е. соlі у 1976 р; потім -для інших видів, в тому числі і для великої рогатої худоби. Також було створено бібліотеки клонів ДНК гіпофіза і гормона росту .
Велике значення мало одержання за допомогою генетичної інженерії інтерферону для людини. Відомо, що інтерферон - це білок, який характеризується універсальною антивірусною дією. Але до останнього часу не була відома амінокислотна послідовність цього білка та не розроблена методика одержання його у чистому вигляді. Тому на першому етапі в крові людини виділили інтерферонову інформаційну РНК, на якій за допомогою ревертази синтезували ген інтерферону. На другому етапі зазначений ген ввели в плазміду і одержали високопродуктивний у-штам бактерій, що виробляв штучний інтерферон. Після того, як була визначена його амінокислотна послідовність та склад нуклеотидів, цей ген було синтезовано хімічним шляхом.
Похожие работы
... . Більш як 40 країн, у тому числі й Україна, підписали Картагенський протокол. Не підписали його лише США, як і Конвенцію про біобезпеку. Треба зазначити, що реакція на продукти з генетично модифікованих джерел їжі різна в США та Європі. Споживачі в США виражають в основному позитивне ставлення до генної інженерії; Під час національного соціологічного опитування, проведеного Міжнародною радою з ...
... до цього, однак різниця в антигенах іноді становиться причиною серйозних захворювань, які несуть загрозу нормальному розвитку вагітності і життю дитини. Частіше всього зустрічається серологічний конфлікт по резус-фактору, який може привести до небезпечних порушень, особливо до гемолітичної хвороби плоду і новонародженого. Кожна людина з моменту свого зачаття має генетично визначену групу крові. І ...

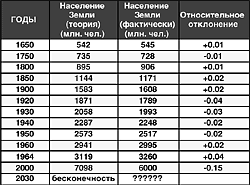

... ЄТЬСЯ, що Одкровення було записано близько 66 року н.е. і, імовірно, доповнене Іоанном згодом через 30 років. З тих пір не проходило жодного століття (а в наш час і жодного року) без нових досліджень і тлумачень цього пророцтва. Число разючих збігів із пророкуванням Іоанна в кожнім столітті було велике, іноді навіть доходило до критичної маси, коли віруючі тієї чи інша країни готувалися до "кінця ...
... дає змогу суттєво розширити виробничі можливості працівників, збільшуючи плідність їх зусиль. У сучасних умовах вона є могутнім фактором розвитку продуктивної сили праці. 4.НАУКОВО-ТЕХНІЧНИЙ ПРОГРЕС ЯК ОСНОВА РОЗВИТКУ ВИРОБНИЦТВА І ЗНИЖЕННЯ ЙОГО ПОТЕНЦІАЛЬНОЇ НЕБЕЗПЕКИ 4.1.Екологічне становище України. Загострення проблем безпеки навколишнього середовнща і населення стало ...
0 комментариев