Навигация
ХАРАКТЕРИСТИКА ОЧИЩЕННЫХ ИНТЕГРАЛЬНЫХ МЕМБРАННЫХ БЕЛКОВ
34143
знака
2
таблицы
2
изображения
3. ХАРАКТЕРИСТИКА ОЧИЩЕННЫХ ИНТЕГРАЛЬНЫХ МЕМБРАННЫХ БЕЛКОВ
Характеристика очищенных мембранных белков, даже самых простых, может составлять определенные трудности. Как и в случае
3.1 МОЛЕКУЛЯРНАЯ МАССА СУБЪЕДИНИЦ
Электрофорез в полиакриламидном геле в присутствии додецил-сульфата натрия — это обычная методика, но в случае интегральных мембранных белков при ее применении возникают особые проблемы. В этом методе додецилсульфат связывается с полипептидными цепями, и комплексы белок—ДНС разделяются в полиакриламидном геле в соответствии с их стоксовыми радиусами, которые в большинстве случаев зависят от молекулярной массы. Молекулярную массу определяют, сравнивая электрофоретическую подвижность данного комплекса и известного стандарта. Однако связывание ДСН с неизвестным белком может качественно отличаться от связывания со стандартами, и тогда будет получен неправильный результат. Подобная ситуация наблюдается для интегральных мембранных белков с высоким содержанием неполярных аминокислотных остатков. С большинством растворимых белков ДСН образует комплексы в соотношении 1,4 г ДСН на 1 г белка, а с белками, содержащими большой процент неполярных остатков, может связываться больше детергента. Возникающий при этом дополнительный отрицательный заряд приводит к аномальному повышению электрофоретической подвижности, и определяемая молекулярная масса оказывается меньше, чем на самом деле. Возможна и другая ситуация. Связывающийся с ДСН мембранный белок может находиться в не полностью развернутом состоянии, что тоже приведет к аномальному повышению электрофоретической подвижности из-за образования более компактного комплекса белок—ДСН. Все эти эффекты весьма существенны. Например, лактозопермеаза имеет кажущуюся мол. массу 33 ООО, если измерять ее с помощью электрофореза в ПААГ в присутствии ДСН; в действительности же, как показывают результаты генетического анализа, ее мол. масса равна 46 ООО. Во многих случаях удается оценить молекулярную массу более точно, если построить график Фергюсона, представляющий собой зависимость электрофоретической подвижности от содержания акриламида как для стандартных белков, так и для исследуемого белка. Этот график зависит от радиуса Стокса и в меньшей степени — от заряда комплекса. Например, по результатам электрофореза в 12%-ном акриламидном геле одна из субъединиц цитохромно-го комплекса Е. coli имеет кажущуюся мол. массу 28 ООО, а из графика Фергюсона получается величина 43 ООО, что совпадает с мол. массой, рассчитанной по данным о секвенировании соответствующей ДНК.
Еще одна проблема — возможное наличие четвертичной структуры. Некоторые мембранные белки агрегируют даже в присутствии ДСН. Например, гликофорин А или белок оболочки бактериофага М13 при электрофорезе в полиакриламидных гелях с ДСН находятся в основном в виде димеров. Иногда агрегация еще более усиливается при нагревании смеси белок—ДСН. Такая картина наблюдается, например, для субъединиц как митохондри-альной, так и бактериальной терминальных оксидаз. Чтобы оценить способность белка к необратимой агрегации, следует провести сравнительный анализ результатов электрофореза в полиакрила-мидном геле с ДСН для прогретых и непрогретых проб. Сходная проблема иногда возникает из-за присутствия детергента, использованного при очистке мембранного белка. Этот детергент необходимо удалить и заменить на ДСН, поскольку в некоторых случаях наблюдается четкая зависимость электрофоретической подвижности от присутствия детергента, с помощью которого солюбилизнровали фермент.
Итак, есть основания думать, что оценка молекулярной массы субъединиц сильно неполярных интегральных мембранных белков, определенная с помощью электрофореза в ПААГ с ДСН, может оказаться неверной. К несчастью, простая альтернатива этому методу отсутствует, и правильную величину часто получают либо по данным о полной первичной последовательности, либо с помощью точного гидродинамического анализа.
3.2 ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ НАТИВНОГО БЕЛКА С ПОМОЩЬЮ ГИДРОДИНАМИЧЕСКИХ МЕТОДОВ
Применение этих методов для мембранных белков может быть сопряжено с большими трудностями, вызванными связыванием детергента. Чтобы оценить это в полной мере, рассмотрим вначале простой растворимый белок, для которого установлена мол. масса субъединиц с помощью электрофореза в ПААГ с ДСН и необходимо узнать, чем он является в неденатурированной, активной форме — мономером, димером или олигомером более высокого порядка. Для определения молекулярной массы белков часто используется гель-фильтрация, включающая сравнение со стандартными белками; здесь возникают проблемы, связанные с тем, что все стандартные белки имеют глобулярную форму, а исследуемый белок может быть не глобулярным, а слегка удлиненным. Такой белок с мол. массой 50 000 может элюировать со скоростью, соответствующей мол. мае
се 100 ООО. В связи с этим колонка для гель-фильтрации должна быть прокалибрована в соответствии со значениями радиуса Стокса, т. е. с размерами «эквивалентной гидродинамической сферы», а кроме того, параллельно необходимо использовать какой-либо другой метод. Обычно измеряют скорость седиментации с помощью либо аналитического ультрацентрнфугирования, либо центрифугирования в градиенте плотности сахарозы. Коэффициент седиментации равен
![]()
где м — молекулярная масса белка,
v — его парциальный удельный объем, ij — вязкость раствора, б — плотность раствора.
Поскольку е и Ч известны, a Rc можно определить с помощью гель-фильтрации, остаются только две неизвестные величины — v и м. Для водорастворимых белков v можно вычислить исходя из аминокислотного состава или непосредственно измерить либо просто принять равным 0,72—0,75 мл/г. Таким образом, измерив S0, можно найти м.
Рассмотрим теперь ситуацию с мембранным белком. Здесь возникают дополнительные проблемы, поскольку гидродинамическая частица — это белково-детергентный комплекс, поэтому м и v в данном случае являются молекулярной массой и удельным объемом комплекса, Мк и К,. К сожалению, К, нельзя оценить, не зная ничего о составе комплекса. В этом случае для нахождения молекулярной массы белка используют два метода.
1.Прямо измеряют количество связанного детергента на 1 г белка. Для этого используют спектральные методы или радиоактивно меченный детергент, а для выделения комплексов применяют различные методы, например гель-фильтрацию. Установив относительное содержание белка и детергента в комплексе, значение К, получают как средневзвешенное соответствующих величин для чистого белка и чистого детергента. После этого без труда находят м„ а поскольку соотношение между белком и детергентом в комплексе известно, находят молекулярную массу белка.
2.Измеряют S0 в средах с разными значениями плотности раствора д. Такие среды обычно получают, используя смеси НгО и D2O. Из графика зависимости S° от q находят как Л/„ так и vt. При этом предполагается, что К, — это средневзвешенное соответствующих величин для чистого белка и чистого детергента.
![]()
ОцеНИВ Квело* и взяв детергент из таблиц, получают молекулярную массу белковой составляющей м,.
Для построения графика зависимости 5° от q проводят аналитическое центрифугирование. Можно проводить центрифугирование и в градиенте плотности сахарозы, используя смеси Н2О и D2O, но анализ результатов в этом случае гораздо сложнее, хотя принципиально не отличается от предыдущего случая.
Альтернативный способ определения молекулярной массы нативной формы мембранного белка состоит в равновесном ультрацентрифугировании. Распределение вещества в состоянии равновесия таково, что наклон графика зависимости логарифма концентрации от г2 равен
где г — расстояние от центра ротора до данной точки в центрифужной пробирке, W — частота вращения.
Если величина У известна или ее легко оценить, как для большинства растворимых белков, эта задача решается достаточно просто. Что касается мембранных белков, то в этом случае определяют на-
Таблица 3. Связывание детергентов с некоторыми мембранными белками
| Белок Na + /К * -АТРаза | Детергент Тритон Х-100 | Количество связанного детергента, мг на 1 мг белка 0,28 | Ссылки |
| Са2+ -АТРаза | Тритон Х-100 | 0,20 | |
| Белок полосы 3 | Тритон Х-100 | 0,77 | |
| Ацетилхолиновый | Тритон Х-100 | 0,70 | |
| рецептор | |||
| Родопсин | Тритон Х-100 | 1,10 | |
| Переносчик | Тритои Х-100 | 1,5 | |
| ADP/ATP | |||
| Инсулииовый | Тритон Х-100 | 0,15; 0,31; 0,54 | |
| рецептор | |||
| Инсулииовый | Дезоксихолат | 0,01; 0,03 | |
| рецептор | |||
| Цитохром | Тритон Х-100 | 0,6 | |
| с-оксидаза | |||
| Цитохром | Лаурилмальтозид | 0,55 |
клон указанной прямой при разных значениях q, получаемых смешиванием НгО и D2O. Как и ранее, одновременно находят Мк и К, и далее определяют молекулярную массу белка.
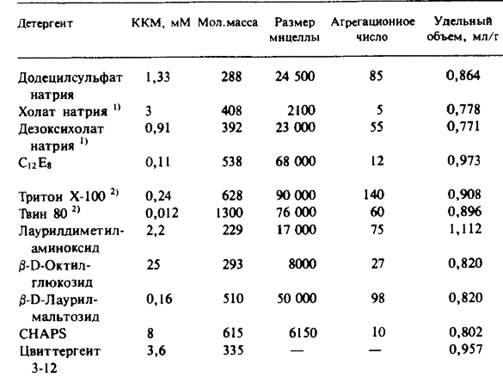
Если в комплексе присутствует третий компонент, возникают дополнительные проблемы. В любом случае все описанные процедуры весьма сложны и могут давать ошибочные результаты. Количество детергента, связанного с очищенными интегральными мембранными белками, может быть весьма существенным — от 0,3 до 1,5 от массы белка, и даже небольшие ошибки в этой величине приведут к значительному искажению молекулярной массы белка. В табл. 3.3 приведены данные о количестве детергентов, присутствующих в некоторых белковых препаратах. Заметим, что растворимые белки с этими детергентами не связываются; это опять свидетельствует о том, что за связывание с детергентом ответственна именно неполярная часть белка, обычно контактирующая с мембранными липидами.
Похожие работы
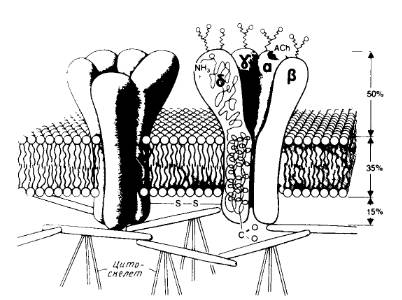
... . О предполагаемом канале, участвующем в переносе, ничего неизвестно; очистка его является довольно сложной задачей. 5. Сборка мультисубъединичных комплексов и обновление мембранных белков После встраивания мембранного полипептида в мембрану он еще должен приобрести правильную конформацию, обеспечивающую его биологическую активность, а если речь идет о мультисубъединичных комплексах, то ...
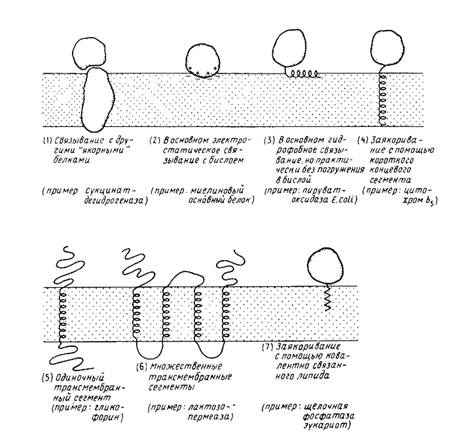
... на N-, либо на С-конце молекулы. N – и С-концы трансмембранных белков (5 и 6) могут находиться как у наружной, так и у внутренней поверхности мембраны. 2. Выделение мембранных белков Очистка и характеристика мембранных белков ставят перед исследователем целый ряд специфических проблем, с которыми он обычно не сталкивается, работая с растворимыми белками. Мембранные белки, как правило, ...
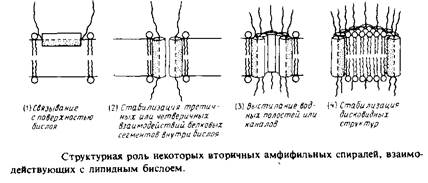
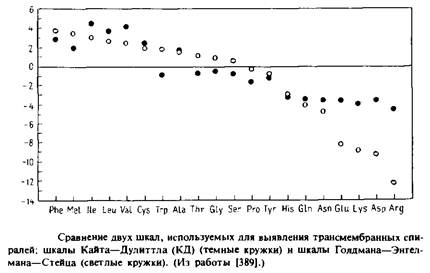
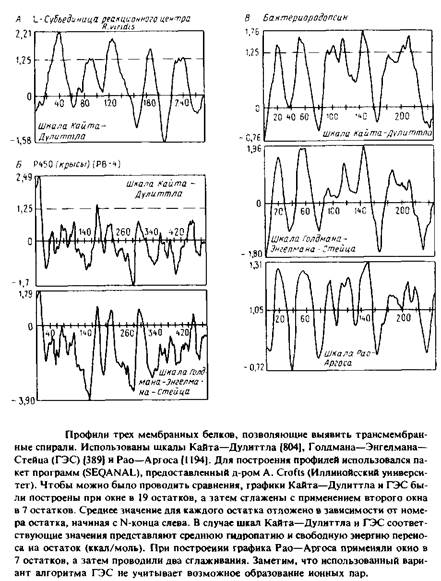
... внутри бислоя могло бы стабилизировать трехмерную структуру мембранных белков. Способы идентификации первичных амфифильных структур Однозначная структурная информация о мембранных белках получена лишь в нескольких случаях, но зато в распоряжении исследователей имеются обширные данные об аминокислотной последовательности, основанные на результатах секвенирования ДНК. Для идентификации ...
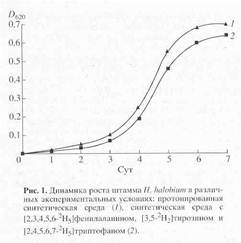
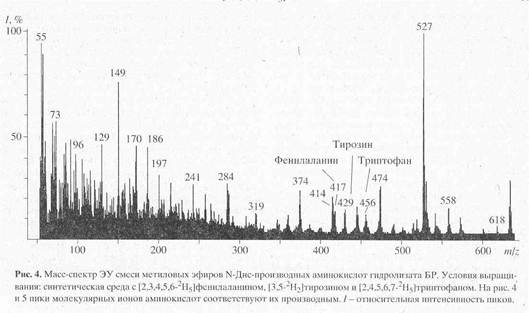
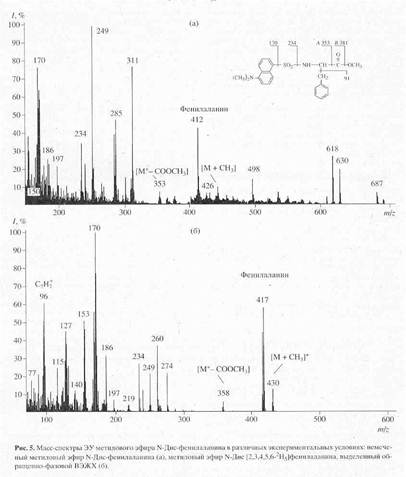
... в его функционировании [10]. Поэтому важно получать подобные модифицированные дейтерием белки в очищенном виде и в препаративных количествах. Целью настоящей работы было получение препаратов бактериородопсина, селективно меченных дейтерием по остаткам ароматических аминокислот - L-фенилаланина, L-тирозина и L-триптофана, а также масс-спектрометрический анализ дейтерий-меченных аминокислот ...
0 комментариев