Навигация
Биосинтез мембранных белков и их встраивание в биомембрану
36107
знаков
0
таблиц
4
изображения
Пензенский Государственный Педагогический Университет
им. В.Г. Белинского
Курсовая работа на тему:
Биосинтез мембранных белков и их встраивание в биомембрану
Выполнила: студентка группы Бх-41
Данилова Елена
Проверил: к.б.н., \
Соловьев В.Б.
Пенза,2009
Содержание
Введение
1. Методы исследования переноса белков через мембраны
2. Встраивание белков в мембрану
2.1 Сигнальная гипотеза
2.2 Мембранная триггерная гипотеза
3. Полипептидные сигналы, отвечающие за сортировку белков и встраивание их в мембраны
3.1 Сигнальная последовательность, определяющая встраивание в эндоплазматический ретикулум
3.2 Стоп-сигналы переноса
3.3 Использование синтетических сигнальных пептидов
3.4 Сигнальные пептидазы
4. Растворимые и мембраносвязанные белки, необходимые для переноса
5. Сборка мультисубъединичных комплексов и обновление мембранных белков
Выводы
Литература
Введение
Процесс образования мембран начинается с синтеза белковых и липидных компонентов, которые затем должны быть доставлены к месту назначения. В состав мембран входят различные белки, для биосинтеза которых необходимы точные механизмы. В принципе имеются две главные проблемы, касающиеся сборки мембранных белков.
1. Все закодированные в ядре белки синтезируются общим пулом рибосом. В связи с этим возникает вопрос: как отдельные мембранные белки доставляются к месту назначения? Чем отличаются белки плазматической мембраны от белков внутренней митохондриальной мембраны или от белков мембран эндоплазматического ретикулума? Эту сложную проблему сортировки можно решить только при наличии определенных сигнальных последовательностей в каждом полипептиде, а также соответствующего аппарата узнавания.
2. Каков истинный механизм встраивания мембранных белков в мембрану и как при этом достигается правильная их ориентация относительно мембранного бислоя? Требуют ли механизмы встраивания и ориентации также наличия определенных сигнальных элементов и систем узнавания и если да, то каковы они? Какие свойства обеспечивают при встраивании мембранных белков формирование правильной третичной, а также четвертичной структуры в случае мультисубъединичных ансамблей?
За последние десятилетия в поиске ответов на эти вопросы достигнуты большие успехи, причем они становятся все более значительными. Это в большой мере обусловлено тем, что для выяснения роли специфических сигнальных полипептидных последовательностей в этих процессах стала использоваться рекомбинантная ДНК. Хотя не на все вопросы удалось найти ответы, полученные результаты все больше убеждают в том, что совершенно разные на первый взгляд системы в действительности обладают фундаментальным сходством. Например, не так давно было показано, что механизм секреции белков имеет много общего с механизмом синтеза белков плазматической мембраны. Совсем недавно достигнут большой прогресс в понимании общих принципов переноса белков через мембраны митохондрий, эндоплазматического ретикулума и грамотрицательных бактерий. Эти экспериментальные системы изучались наиболее интенсивно. И хотя между соответствующими процессами есть значительные различия, они имеют ряд общих особенностей.
1. Существует идентифицируемая часть полипептидной последовательности, которая служит участком узнавания, или «сигналом», направляющим отдельный полипептид к мембране, в которую он встраивается. Эти сигнальные участки часто расположены на N-конце новосинтезированного полипептида и отщепляются специфическими сигнальными пептидазами после встраивания его в нужную мембрану или переноса через нее. Для обозначения N-концевого сигнала различными авторами использовались следующие термины: сигнальный пептид, сигнальная последовательность, транзитный пептид, лидирующий пептид пре-последовательность.
2. Процессы трансляции и встраивания белков в мембрану можно разделить в эксперименте. Для сборки мембранных белков в большинстве случаев необходима энергия, отличающаяся по величине от той, которая требуется для их трансляции на рибосоме.
3. Связавшийся с мембраной-мишенью полипептид должен, кроме того, находиться в конформации, в которой может осуществляться его перенос через мембрану или встраивание в нее. Во многих случаях перенос белков через мембраны происходит от N-конца к С-концу, при этом необходимо, чтобы белок был, по крайней мере, частично развернут или слабо свернут. Полипептид может транслоцироваться в вытянутой форме в ходе энергозависимого процесса.
Особый интерес представляет процесс сборки мембранных белков. На рис.1 схематично показаны три общих механизма проникновения пептидного предшественника в мембрану. Механизмы А и Б являются вариантами схемы линейного вытеснения, согласно которой сигнальная последовательность направляет полипептид к переносящему устройству, которое включает в себя заполненный водой канал. Сигнальная последовательность может проходить прямо сквозь канал (механизм А) или оставаться связанной с мембраной, образуя, как показано на рис.1, петлю (механизм Б). В отсутствии какого-либо сигнала остановки процесса переноса полипептид будет транспортироваться через мембрану целиком. Однако если внутри полипептида имеется второй сигнальный пептид, называемый стоп-сигналом переноса, то процесс останавливается, и стоп-сигнал переноса становится трансмембранным сегментом зрелого мембранного белка. Фиксируя белок в мембране, стоп-сигнал переноса действует как сигнал сортировки.
Схема В на рис.1 иллюстрирует возможную роль самопроизвольного включения в мембрану гидрофобных элементов полипептидного предшественника. Этот механизм может реализовываться только тогда, когда включение в мембрану происходит после трансляции полипептида.
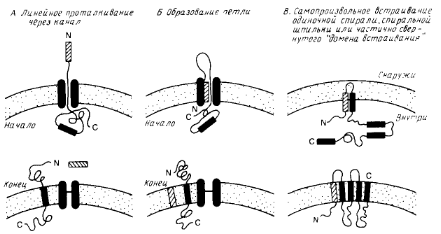
Рис.1. Три общие модели возможной сборки белков в мембране.
Две первые (А и Б) предполагают, что белок транспортируется в линейной форме через белковый канал. При наличии стоп-сигнала процесс останавливается, в противном случае через мембрану проходит весь белок. Модель В предполагает, что гидрофобные элементы полипептида самопроизвольно включаются в липидный бислой. Гидрофобный элемент может представлять собой одиночную спираль или более сложную структуру. Процесс может быть опосредован белками.
Проблема сборки белков очень важна. Этот процесс обычно не протекает самопроизвольно, лишь в результате взаимодействия между образующимися полипептидами и липидным бислоем. Напротив, он является энергозависимым и опосредуется белковыми структурами, которые пока не изучены в достаточной степени. Экспериментальные данные свидетельствуют о том, что перенос белков через мембрану (например, в полость эндоплазматического ретикулума) и сборка интегральных мембранных белков – это тесно связанные стороны одного и того же процесса. Логично ожидать, что проблемы транспортировки белков через мембраны и их укладки должны решаться одинаковым образом.
Похожие работы
... . Все четыре системы являются основными белковыми компонентами мембран, в которых они локализованы, и могут служить иллюстрацией различных уровней организации мультиферментных комплексов в мембранной энзимологии. Первые две системы катализируют анаболические и катаболические реакции, протекающие в присутствии молекулярного кислорода и обычно липофильных мембраносвязанных субстратов. Терминальные ...
... инженерию. Необходимо отметить, что если базовый стандарт по химии не предусматривает изучение вопросов биотехнологии, то таковой по биологии содержит наиболее общие её аспекты: достижения генной инженерии и перспективы биотехнологии. 2.2 Межпредметные связи по изучению аспектов биотехнологии в средней школе По программе Р.Г. Ивановой и Л.А. Цветкова в 10 классе предусмотрено изучение темы ...
... векторов для экспрессии. 2.2 Способы прямого введения гена в клетку Прямое введение гена в клетку осуществляют несколькими способами: 1. Трансфекция 2. Микроинъекция 3. Электропорация 4. Метод «мини-клеток» 5. Упаковка в липосомы 6. Электронная пушка При трансфекции ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, ...
0 комментариев