Навигация
1.2 Протеинкиназа
Через 10 лет после открытия Сазерлендом цАМФ была обнаружена цАМФ-зависимая протеинкиназа, значительно повышающая скорость фосфорилирования субстратов в присутствии цАМФ. Реакция протеинфосфорилирования выглядит следующим образом:
![]()
Гидроксильная группа белка, акцептирующая терминальный фосфат АТФ, почти всегда принадлежит серину, реже – треонину и тирозину. Фосфосериновые остатки в белках существенно ионизированы при физиологических значениях рН, поэтому фосфорилирование или дефосфорилирование серина резко меняет заряд белковых молекул. После открытия протеинкиназы А стало ясно, что реакции фосфорилирования – дефосфорилирования опосредуют действие многих гормонов и нейромедиаторов, которые через р-адренергическую рецептию активируют аденилатциклазу и приводят к повышению внутриклеточного уровня цАМФ.
Протеинкиназа А обнаружена во всех нормальных клетках млекопитающих, а также в некоторых типах клеток немлекопитающих. Высоким уровнем протеинкиназы А отличается мозг, где фермент распределен равномерно по всем отделам. Для проявления активности фермента необходимы ионы магния. Так, из легких кролика выделен стимулирующий модулятор протеинкиназы, связывающий магний и названный "магмодулином". Протеинкиназа А – тетрамер, построенный из двух различных субъединиц: регуляторной, связывающей цАМФ, и каталитической, осуществляющей фосфотрансферазную реакцию; будучи соединены вместе, они образуют неактивный комплекс. При активации происходит связывание цАМФ Р-субъединицей холофермента, после чего возможна диссоциация Р от К-субъединицы. Свободная К-рубъединица способна катализировать фосфорилирование различных клеточных белков:
![]()
Таким образом, в активном состоянии протеинкиназа А представляет димер Р-субъединиц, связанный с 4 молями цАМФ, и 2 свободные К-субъединицы.
Скорость обратной реакции обусловлена не только спонтанной диссоциацией комплексов Р – цАМФ, но и концентрацией К-субъединиц, так как последние высвобождают связанный цАМФ путем рекомбинации с комплексом Р – цАМФ. Этот своеобразный "ретрокооперативный" эффект К-субъединицы, названный так Свилленсом и Дюмоном, ведет к временному увеличению концентрации цАМФ, требуемой для активации протеинкиназы. В период стимуляции системы уровень К-субъединиц резко падает.
Существование эквивалентных количеств каталитических и регуляторных субъединиц цАМФ-зависимой протеинкиназы во многих тканях привело к представлению о том, что единственная функция регуляторной субъединицы – контроль протеинкиназной активности. Однако регуляторная субъединица может существовать отдельно от каталитической в некоторых типах клеток, например в клетках нейробластомы. Это указывает на возможность проявления биологического действия регуляторной субъединицы независимо от каталитической. Далее будет рассмотрена такая возможность на примере изменения проницаемости мембран нейронов для Na+ и К в присутствии Р-субъединицы. Надо также иметь в виду, что цАМФ-связывающие белки, не ассоциированные с протеинкиназой, могут регулировать активность последней путем модулирования уровня свободного циклического нуклеотида, способного присоединяться к протеинкиназе.
Отметим далее, что свободные Р-субъединицы ингибируют фосфодиэстеразу цАМФ, что может приводить к увеличению уровня этого нуклеотида в клетке. Установлено также, что свободные Р-субъединицы протеинкиназы типа А в отличие от холофермента киназы и ее К-субъединицы ингибируют фосфопротеинфосфатазы нескольких типов. Ингибирование обусловлено снижением скорости катализа без изменения сродства к энзиму. Очевидна физиологическая значимость такого ингибирования, так как в этом случае диссоциация А-киназы может способствовать не только стимулированию фосфорилирующей активности, но и ингибированию дефосфорилирования субстратов.
Протеинкиназа А существует в форме двух изоферментов, относительное количество которых варьирует в разных тканях.
Изоферменты были названы киназами I и II. Они имеют различные скорости диссоциации в присутствии гистона и растворов NaCl и реассоциации после удаления цАМФ. Так, киназа I типа быстро диссоциирует в присутствии гистона или 0,5 М NaCl и медленно реассоциирует после удаления цАМФ; напротив, киназа II типа медленно диссоциирует в присутствии указанных агентов и быстро реассоциирует после удаления цАМФ.
Каталитические субъединицы киназ I и II типа имеют молекулярную массу 40 кД и минимальные различия в аминокислотном составе. Напротив, регуляторные субъединицы киназ I и II типа значительно отличаются по первичной структуре. Вероятно, субъединица Р I типа – 49 кД – является протеолитическим фрагментом Р II типа с молекулярной массой 55 кД. Один из сериновых остатков Р-субъединицы II типа фосфорилируется каталитической субъединицей цАМФ-зависимой протеинкиназы. Очищенная же Р-субъединица I типа не фосфорилируется каталитической субъединицей цАМФ-зависимой протеинкиназы. Другим отличием киназы I типа от II типа является то, что только первый фермент связывает Mg-АТФ с высоким сродством.
цАМФ-зависимые протеинкиназы локализованы в основном в цитозольной фракции клеток. Однако в мозге, например, значительная часть киназы II типа является мембранно-связанной. Очевидно, субклеточная локализация и соотношение киназ I и II типа могут обусловливать специфику действия цАМФ в клетке. Отношение киназы I типа к киназе II типа варьирует в различных органах и в различных фазах клеточного цикла.
При исследовании локализации фосфорилирующих систем в ЦНС установлено, что цАМФ-зависимая система избирательно сконцентрирована в нейронах, особенно в дендритах, а не в глии. Мозг крысы содержит как I, так и и II форму протеинкиназы А при соотношении этих форм 1:4 соответственно. Высокое содержание А-киназы II типа по сравнению с ферментом I типа вообще характерно для нервной ткани. Недавно установлена гетерогенность Р-субъединиц А-киназы II типа. При этом в мозге выявлена собственная, специфичная Р Н-субъединица, отличная по иммунохимическим свойствам от Р-субъединиц II типа в других тканях. Р II мозга отличается от Р II – мышц по характеру взаимодействия с К-субъединицей, а также по электрофоретической подвижности аутофосфорилированных форм. Показано, что фракция Р-субъединиц II типа мозга взаимодействует с Са+ и кальмодулином. Необычные свойства Р Н-субъединицы мозга могут являться следствием адаптации нервных клеток к специфической функции передачи и хранения информации.
Ассоциации А-киназы II типа мозга с мембранной фракцией клеток обусловлены только Р-субъединицей. цАМФ при диссоциации холофермента высвобождает из мембранно-связанного состояния только К-субъединицу; таким образом, компартментализация К-субъединицы изменяется при активации А-киназы. Молекула этой субъединицы гидрофильна, поэтому диссоциация мембранно-связанной А-киназы II типа приводит к транслокации каталитической субъединицы в цитоплазму и далее в ядро. Предполагается, что такая транслокация может обусловливать цАМФ-зависимое изменение экспрессии генов в нейронах, наибольшая концентрация Р-субъединиц А-киназы II типа обнаружена в пре- и постсинаптической мембранах, что свидетельствует о важной роли этой киназы в синаптической передаче.
Различия между регуляторными субъединицами А-киназы II типа нервной и других тканей проявляются также и во взаимодействии этих субъединиц с субстратами киназы. Так, для Р-субъединицы II типа из мозга характерно тесное взаимодействие с МАР-2 – нейроспецифическим белком, локализованным в отростках нейронов, кальцинейрином и другими белками. Такое взаимодействие Р II с субстратами А-киназы может приводить, с одной стороны, к локализации А-киназы II типа около специфических субстратов и, соответственно, в определенных внутриклеточных компартментах, что, очевидно, имеет важное физиологическое значение. С другой стороны, кроме связывания К-субъединицы Р-субъединица II типа нервной ткани может участвовать в регуляции функционирования других белков.
Регуляция активности протеинкиназы А осуществляется несколькими путями. Так, во многих тканях, в том числе и нервной, обнаружен низкомолекулярный термостабильный ингибитор киназы. Ингибитор связывается со свободной К-субъединицей и угнетает ее ферментативную активность. Вероятно, физиологическая роль ингибитора состоит в блокаде фосфорилирующей активности при "базаяном", т.е. нестимулируемом уровне цАМФ. В мозге наблюдается изменение концентрации ингибитора в ответ на некоторые гормональные сигналы, что может иметь определенное значение в долговременной регуляции активности А-киназы. Регуляция активности киназы II типа осуществляется также аутофосфорилированием ее Р-субъединицы; это ведет к повышению активности фермента, что обусловлено уменьшенной скоростью реассоциации фосфорилированной Р со свободной К-субъединицей. Киназа I типа также может быть фосфорилирована, но фосфорилирование ее Р-субъединицы осуществляется цГМФ-зависимой протеинкиназой. Установлено ау-тофосфорилирование К-субъединицы, однако функциональное значение этого процесса неизвестно.
Возможно, что фосфорилирование с помощью свободной К-субъединицы А-киназы и соответствующая активация фосфопротеинфосфатазного ингибитора I, приводящая к снижению активности фосфопротеинфосфатазы I, уменьшает дефосфорилирование комплекса Р-цАЛ1Ф и, следовательно, способствует диссоциации киназы II типа по принципу положительной обратной связи. Такое событие наряду с компартментализацией цАМФ в клетке может способствовать эффективной активации цАМФ-зависимой киназы при незначительном увеличении внутриклеточного уровня цАМФ.
цАМФ-зависимое фосфорилирование белков нервных окончаний оказывает существенное влияние на синаптическую передачу. Так, установлено, что мутации у дрозофил, изменяющие метаболизм цАМФ, приводят к плейотропным нарушениям механизма обучения. У моллюска аплизии серотонинергическая синаптическая передача усиливается инъекцией цАМФ в нейрон или инкубацией ганглия с блокаторами фосфодиэстеразы. Так как известно, что входящий ток кальция является непременным посредником в осуществлении синаптического действия, полагают, что серотонин усиливает такой ток посредством повышения внутриклеточного уровня цАМФ и активности А-киназы. Прямые доказательства осуществления этого механизма получены в экспериментах, где отмечено увеличение Са-проводимости нейронов улитки при инъекции в них К-субъединицы А-киназы. Блокаторы протеинкиназы А оказывают противоположный эффект – быстрое снижение Са-проводимости.
Повышение уровня ионизированного Са+ внутри клетки при открывании Са-каналов может способствовать снижению в ней содержания цАМФ за счет активации Са – КМ-зависимой фосфодиэстеразы и по принципу отрицательной обратной связи приводить к переходу Са-каналов в неактивное состояние. Введение в нейрон теофиллина замедляет процесс ослабления Са-проводимости. Таким же влиянием на кальциевую проводимость обладают ингибиторы кальмодулина, например трифтазин.
Таким образом, функционирование Са-каналов прямо регулируется протеинкиназой А, осуществляющей фосфорилирование их белковых компонентов, а также опосредованно контролируется гормонами, медиаторами и другими факторами, вызывающими или активацию аденилатциклазы, или ее ингибирование. При нарушении фосфорилирования каналы быстро теряют способность активироваться под действием изменения трансмембранного электрического поля. В течение некоторого времени это состояние является обратимым и фосфорилирование снова переводит каналы в рабочее состояние либо изменяя кинетику их активации, либо включая ранее неактивные каналы. При длительном нарушении цАМФ-зависимого фосфорилирования каналы подвергаются необратимым изменениям.
Дополнительный путь для токов Са+ внутрь клетки могут образовывать хемоуправляемые каналы, непосредственно активируемые медиаторными веществами в ответ на их взаимодействие с рецепторами. В отличие от электроуправляемых Са-каналов время жизни этих каналов существенно больше, однако относительное количество ионов, переносимых через хемоуправляемые каналы, вероятно, невелико по сравнению с электроуправляемым путем. По ряду данных хемоуправляемые каналы полностью инактивируются при увеличении внутриклеточного содержания цАМФ.
Заслуживает также рассмотрения участие цАМФ в К- и Na-проводимости мембраны.
Известно, что болевое раздражение усиливает рефлекс втягивания жабры у моллюска эплизии за счет модуляции секреции передатчиков сенсорными нейронами. Повышение чувствительности происходит за счет снижения К-проводимости и соответствующего увеличения продолжительности потенциала действия, в результате чего усиливаются Са-рефлекс и экзоцитоз. Болевое раздражение вызывает выделение серотонина "облегчающим" нейроном в окончаниях сенсорных нейронов. Серотонин, в свою очередь, увеличивает синтез цАМФ, активность протеинкиназы А и степень фосфорилирования белка, тесно связанного с К-каналом. В результате происходит закрытие канала. Блокада К-каналов приводит к тому, что приходящий в нервное окончание потенциал действия спадает медленно: "продленные" потенциалы действия удерживают потенциал-зависимые Са-каналы в открытом состоянии, вследствие чего приток ионов Са возрастает. Это, в свою очередь, ведет к опорожнению большего числа синаптических пузырьков. К блокаде К-каналов приводит также добавление АТФ и К-субъединицы протеинкиназы А в мембранные препараты нейронов; добавление протеинфосфатазы обусловливает открывание К-каналов.
Электрозависимые Na-каналы ответственны за формирование потенциала действия в возбудимых мембранах. Установлено, что в синаптосомах мозга а-субъединица №^каналов, играюшая центральную роль в функционировании этих каналов, фосфорилируется как эндогенной, так и экзогенной протеинкиназой А. Фосфорилирование приводит к ингибированию активированного нейротоксинами входящего Na-тока. Таким образом, активность Са-каналов, К-каналов и Na-каналов нейронов модулируется под действием протеинкиназы А и / или через изменение скорости их дефосфорилирования протеинфосфатазами.
Отметим, что протеинкиназа А в нервной ткани регулирует также чувствительность р-адренорецепторов к агонистам. Так, десенситизация этих рецепторов коррелирует с их цАМФ-зависимым фосфорилированием. Установлена также регуляция А-киназой биосинтеза самих р-агонистов. Эта регуляция осуществляется с помощью цАМФ-зависимого фосфорилирования тирозингидроксилазы – узлового фермента биосинтеза катехоламинов. Такое фосфорилирование может быть составной частью механизма ускорения биосинтеза катехоламинов в ответ на нервный импульс или секрецию нейромедиаторов в нервной ткани в условиях in vivo,
Остановимся на роли цАМФ и еще одного вторичного посредника – 2', 5'-олигоаденилата в регуляции пролиферации и дифференцировки нервных клеток. 2–5А – это олигонуклеотид, состоящий из нескольких остатков АМФ, связанных 5'-фосфодиэфирной связью. В клетке существуют 2 фермента, обеспечивающие определенный уровень 2–5А. Олигосинтетаза – фермент биосинтеза 2–5А, активен лишь в присутствии двуспиральной РНК; молекулярная масса – 100–105 кД. 2'-Фосфодиэстераза гкдролизует 2–5А до АМФ и АТФ, молекулярная масса этого фермента около 40 кД.
Чтобы оценить биохимическое и физиологическое значение процессов образования 2–5А, необходимо остановиться на механизмах подавляющего влияния цАМФ на пролиферативный статус нервных клеток. Так, действие ряда факторов, в частности фактора роста нервов, а также повышение уровня цАМФ, вызванное теофиллином или дибутирил-цАМФ, приводит к остановке деления и дифференцировке этих клеток. Действие теофиллина в отличие от фактора роста нервов или дибутирильного аналога цАМФ носит кратковременный характер. Повышение концентрации цАМФ в результате ингибирования фосфодиэстеразы под действием теофиллина, как и в случае фактора роста нервов и дибутирил-цАМФ, приводит к блокаде активного деления и дифференцировке нервных клеток. Однако в дальнейшем падение активности фосфодиэстеразы, обусловленное теофиллином, и соответствующее увеличение уровня цАМФ компенсируется индукцией биосинтеза этого фермента. Именно этим и объясняется кратковременное действие теофиллина.
Повышение уровня цАМФ в нервных клетках, вызванное действием факторов, непосредственно влияющих на аденилатциклазную систему, сопровождается более чем 10-кратным увеличением концентрации 2–5А. Установлено, что рост уровня олиго синтетазы и концентрации 2–5А наблюдается при дифференцировке клеток и замедлении клеточного деления.
Исследование биологических свойств 2–5А показало, что этот олигоиуклеид способен обратимо активировать специфическую латентную нуклеазу, ускорять гидролиз РНК и, таким образом, ингибировать синтез белка in vivo и тормозить процесс размножения клеток. Следовательно, повышение уровня 2–5А и изменение концентрации цАМФ является частью универсального механизма регуляции клеточного деления. К настоящему времени установлены два взаимодополняющих процесса, с помощью которых осуществляются эти регуляторные реакции. С одной стороны, стимуляция А-киназы при повышении уровня цАМФ приводит к фосфорилированию белка сМг – 18 кД. Этот белок в виде фосфоформы является ингибитором активности 2' – фосфодиэстеразы – фермента гидролиза 2–5А. С другой стороны, повышение концентрации цАМФ внутри клетки вызывает индукцию олиго синтетазы. осуществляющей синтез 2–5А.
Совокупность этих событий приводит к устойчивому повышению уровня 2–5А и связанному с этим переходу нервных клеток в состояние покоя. По-видимому, именно таким образом осуществляется ярко выраженный антинролиферативный эффект цАМФ, вызванный действием этого вторичного посредника на систему метаболизма 2–5А. При этом взаимное влияние уровней цАМФ и 2–5А основано на механизме отрицательной обратной связи. Так, установлено, что подъем уровня 2–5А, обусловленный увеличением внутриклеточной концентрации цАМФ по указанным механизмам, в свою очередь, способствует активации фосфодиэстеразы цАМФ. В результате происходит снижение уровня цАМФ и прекращение роста концентрации 2–5А, который является в этом случае вторичным посредником в проявлении антипролифера-тивного действия цАМФ.
Похожие работы
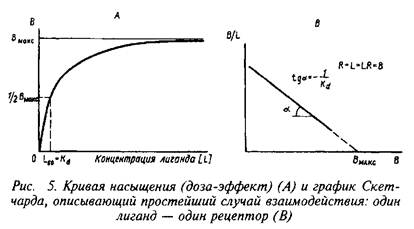
... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
... мышление и др., а также индивидуальные особенности (способности). Таким образом, проблема научения является одной из фундаментальных в психологии вообще и психофизиологии в частности, поскольку она позволяет понять психическую адаптацию человека к условиям существования, сколь бы они ни были необычными, своеобразными, чрезвычайными. В непосредственной связи с научением находится проблема памяти. ...
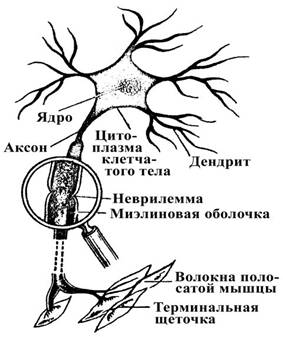
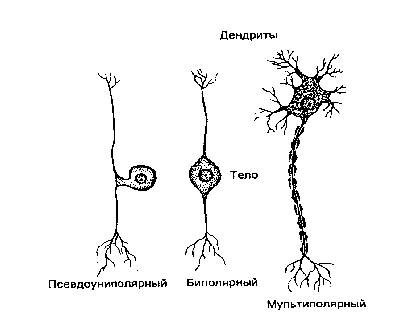
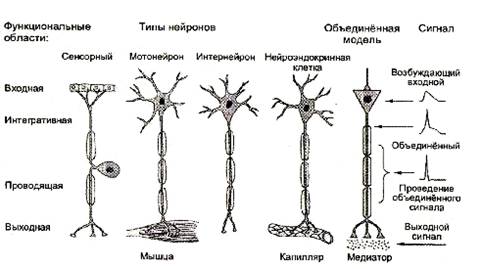
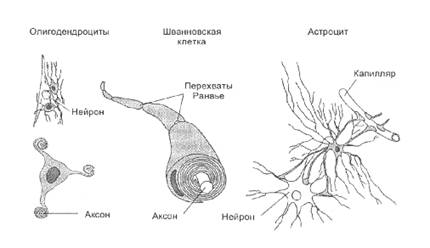
... плотно смыкаются друг с другом и практически полностью обвертывают кровеносный сосуд. Подобная изоляция сосуда является одним из способов формирования гематоэнцефалического барьера - граници между кровью и нервной тканью, закрытой для многих находящихся в крови веществ. Другие отростки астроцита почти целиком обвертывают тела нейронов. Если нейрон возбуждается длительно, вокруг него повышается ...
... . И.П. Павлову, первому из русских ученых, 7 октября 1904 г. была присуждена Нобелевская премия в знак признания его работ по физиологии пищеварения. Потребность организма в пище проявляется в виде физиологической реакции голода. У человека голод приобретает выраженную субъективную окраску — от относительного безразличия к пище до яркой эмоциональной реакции. Физиологической основой голода ...
0 комментариев