Навигация
Белоксинтезирующая система
26238
знаков
2
таблицы
4
изображения
2. Белоксинтезирующая система
Клетки должны обладать специальными механизмами для точного, аккуратного и эффективного перевода последовательности мРНК в соответствующую последовательность аминокислот кодируемого белка. Трансляция (биосинтез белков с использованием мРНК в качестве матрицы) осуществляется в клетках при помощи сложной белок-синтезирующей системы. Отдельные компоненты этой системы ассоциируют в единую структуру по мере ее функционирования и разобщаются по окончанию синтеза. В состав белок-синтезирующей системы входят следующие структуры:
· Рибосомы;
· матричная РНК;
· транспортная РНК;
· белковые факторы и ферменты инициации, элонгации и терминации трансляции;
· набор аминокислот;
· набор аминоацил-тРНК-синтетаз, образующих аминоацил-тРНК;
· макроэрги АТФ и ГТФ;
· ионы Mg 2+, Ca2+, K+, NH4+.
РибосомыРибосомы представляют собой рибонуклеопротеиновые образования — своеобразные "фабрики", на которых идёт сборка аминокислот в белки. Обычно рибосомы характеризуют по скорости их седиментации в центрифужном поле, которая количественно выражается константой седиментации s, выражаемой в единицах Сведберга S. Эукариотические рибосомы имеют константу седиментации 80S и состоят из 40S (малой) и 60S (большой) субъединиц. Каждая субъединица включает рРНК и белки. В 40S субъединицу входит рРНК с константой седиментации 18S и около 30—40 белков. В 60S субъединице обнаружено 3 вида рРНК: 5S, 5,8S и 28S и около 50 различных белков.
Белки входят в состав субъединиц рибосомы в количестве одной копии и выполняют структурную функцию, обеспечивая взаимодействие между мРНК и тРНК, связанными с аминокислотой или пептидом.
В присутствии мРНК 40S и 60S субъединиц объединяются с образованием полной рибосомы, масса которой примерно в 650 раз больше массы молекулы гемоглобина.
В рибосоме есть 2 центра для присоединений молекул тРНК: аминоацильный (А) и пептидильный (Р) центры, в образовании которым участвуют обе субъединицы. Вместе центры А и Р включают участок мРНК, равный 2 кодонам. В ходе трансляции центр А связываем аа-тРНК, строение которой определяет кодон, находящийся в области этого центра. В струкЯ туре этого кодона зашифрована природа аминокислоты, которая будет включена в растущую полипептидную цепь. Центр Р занимает пептидил-тРНК, т.е. тРНК, связанная с пептидной цепочкой, которая уже синтезирована.
У эукариотов различают рибосомы 2 типом "свободные", обнаруживаемые в цитоплазма клеток, и связанные с эндоплазматическим ретикулумом (ЭР). Рибосомы, ассоциированнье с ЭР, ответственны за синтез белков "на экспорт", которые выходят в плазму крови и участвуют в обновлении белков ЭР; мембраны аппарата Гольджи, митохондрий или лизосом.
Митохондрии содержат свой набор рибосом. Митохондриальные рибосомы мельче, чем рибосомы эукариотов, прокариотов и имеют константу седиментации 55S. Они также состояв из двух субъединиц, но отличаются от эукаририотических рибосом количеством и составом РНК и белков.
Матричная РНК
Содержит информацию о структуре синтезируемого белка и используется в качестве матрицы.
Опыты Ниренберга свидетельствуют о том, что не рибосома и не рРНК являются матрицей, на которой синтезируются специфические белки, эту роль выполняют поступающие извне матричные РНК. ДНК предает информацию на РНК, которая синтезируется в ядре и затем поступает в цитоплазму. Здесь РНК выполняет матричную функцию для синтеза специфической белковой молекулы. Матричная гипотеза синтеза белка, как и других полимерных молекул ДНК и РНК, получила в настоящее время полное подтверждение. Ее правильность была доказана в экспериментах, которые обеспечивали точное воспроизведение первичной структуры полимерных молекул; причем этот синтез в отличии от беспорядочного химического синтеза отличался не только высокой скоростью и специфичностью, но и направленностью самого процесса, в строгом соответствии с программой, записанной в линейной последовательности молекулы матрицы.
Транспортная РНК
На долю тРНК приходится около 10-15% общего количества клеточной РНК. К настоящему времени открыто более 60 различных тРНК. тРНК называют "адапторные молекулы", т.к. к акцепторному концу этих молекул может быть присоединена определенная аминокислота, а с помощью антикодона они узнают специфический кодон на мРНК. Для каждой аминокислоты в клетке имеется по крайней мере одна специфическая РНК (для ряда аминокислот открыто более одной, в частности для серина – 5 разных тРНК, для лизина и глицина – по 4 разных тРНК, хотя и в этом случае каждая тРНК связана со специфической аминоацил–тРНК-синтетазой). Молекулярная масса большинства тРНК колеблется от 24000 до 29000 Да. Они содержат от 75 до 85 нуклеотидов. Аминокислоты присоединяются к свободной 3¢-ОН-группе концевого мононуклеотида, представленного во всех тРНК АМФ, путем образования эфирной связи. Интересно, что почти все тРНК обладают не только индивидуально сходными функциями, но и очень похожей трехмерной структурой.
Установлена первичная структура почти всех 60 открытых тРНК. Общей для тРНК оказалась также нативная конформация, установленная методом рентгеноструктурного анализа и названная первоначально названная конформацией клеверного листа; на самом деле эта конформация имеет неправильную, Г-образную, форму.
Определение структуры тРНК позволило выявить ряд отличительных участков; так, 3¢-гидроксильном конце располагается одинаковая для всех тРНК последовательность триплета ЦЦА – ОН, к которой присоединяется посредством эфирной связи специфическая аминокислота. Связывание в основном происходит через 3¢-ОН-группу концевого аденилового нуклеотида, хотя получены доказательства возможности присоединения аминокислоты и через 2¢-ОН-группу. Тимидин-псевдоуридин-цитидиловая петля, по-видимому, обеспечивает связывание аминоацил-тРНК с поверхностью рибосомы. Имеется кроме того, добавочная петля, состав которой варьирует у разных типов молекул тРНК; ее назначение неизвестно. Дигидроуридиловая петля, с другой стороны, оказалась необходимой как сайт (место) для узнавания специфическим ферментом – аминоацил-тРНК-синтетазой. Имеется также антикодоновая петля, несущая триплет, названный антикодоном, и расположенная на противоположной стороне от того конца, куда присоединяется аминокислота. Антикодон является антипараллельными в своей комплементарности.
Белковые факторы
В каждой стадии белкового синтеза на рибосоме: инициации, элонгации и терминации участвует разный набор внерибосомных белковый факторов:
- инициации elF1, elF2;
- элонгации FF1, FF2;
- терминации RF1, RF2, RF3;
- другие.
Эти белки связываются с рибосомой или её субъединицами на определённых стадиях процесса и стабилизируют или облегчают функционирование белоксинтезирующей машины.
Аминокислоты
Все 20 аминокислот, входящих в структуру белков организма человека, должны присутствовать в достаточном количестве. Это требование прежде всего относится к незаменимым (т. е. не синтезирующимся в организме) аминокислотам, так как недостаточное снабжение клетки хотя бы одной незаменимой аминокислотой приводит к снижению, а иногда и полной остановке синтеза белка на кодоне, требующем включения этой аминокислоты в белок.
Аминоацил –тРНК синтетазыТак как у нуклеиновых кислот нет какого-либо специального сродства к боковым цепям аминокислот, взаимное узнавание должно происходить с помощью специальной молекулы белка, способной выявлять одновременно и определенную тРНК-молекулу, и соответствующую аминокислоту. Для подобного узнавания и правильного присоединения соответствующей аминокислоты к молекуле тРНК должно существовать по крайней мере 20 специфичных ферментов. Процесс узнавания и присоединения происходит в два этапа и катализируется ферментом – уникальным для каждой из 20 аминокислот, принадлежащим к классу аминоацил-тРНК-синтетаз. Этот фермент образует активированный промежуточный аминоацил-АМР-ферментативный коплекс, который специфически узнает соответствующую молекулу тРНК и переносит аминокислотный остаток на 3¢-OH группу концевого аденозина. Аминокислота остается присоединенной эфирной связью к тРНК вплоть до включения в определенное положение растущей полипептидной цепи предшественника белка.
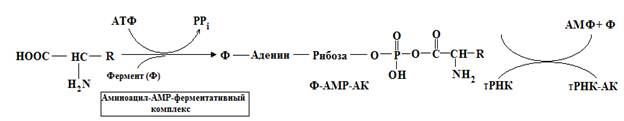
На включение одной аминокислоты в растущую полипептидную цепь клетка затрачивает 4 макроэргические связи: 2 из АТФ в ходе реакции, катализируемой аа-тРНК синтетазой (в процессе активации аминокислот АТФ расщепляется на АМФ и пирофосфат), и 2 молекулы ГТФ: одна используется на связывание аминоацил-тРНК в А-центре рибосомы, а вторая затрачивается на стадию транслокации. К этому следует добавить использование ещё двух макроэргических связей молекул: АТФ и ГТФ на инициацию и терминацию синтеза полипептидной цепи.
3. Этапы синтеза полипептидной цепиСинтез белка представляет собой циклический многоступенчатый энергозависимый процесс, в котором свободные аминокислоты полимеризуется в генетически детерминированную последовательность с образованием полипептидов. Система белкового синтеза, точнее система трансляции, которая использует генетическую информацию, транскрибированную в мРНК, для синтеза полипептидной цепи с определенной первичной структурой, включает около 200 типов макромолекул – белков и нуклеиновых кислот. Среди них около 100 макромолекул, участвующих в активировании аминокислот и их переносе на рибосомы, более 60 макромолекул, входящих в состав 70S или 80S рибосом, и около 10S макромолекул, принимающих непосредственное участие в системе трансляции. Белковый синтез, или процесс трансляции, может быть условно разделен на 2 этапа: активирование аминокислот и собственно процесс трансляции.
Второй этап матричного синтеза белка, собственно трансляцию, протекающей в рибосоме, условно делят на три стадии: инициации, элонгации и терминации.
Активирование аминокислот
Большая часть аминокислот в цитоплазме клеток находиться не в свободном состоянии, а в виде аминоацил-тРНК. Это предохраняет аминокислоты от метаболических превращений и способствует сохранению набора аминокислот для синтеза белка. Образованию комплекса аминокислота-тРНК предшествует активация аминокислоты и нахождение соответствующей тРНК (рекогниция). Это происходит под действием фермента аминоацил-тРНК-синтетазы, или АРС-азы. Эти ферменты имеют два активных центра, один из которых соответствует определенной тРНК, а другой строго специфичен соответствующей аминокислоте. Таким образом, в клетке должно быть не менее 20 АРС-аз, хотя фактически их несколько больше. Образование аминоацил-тРНК происходит в два этапа, первым из которых является взаимодействие АК с АТФ:
![]()
Аминоациладенилат (АК~АМФ) остается в комплексе с АРС-азой до присоединения ко второму активному центру фермента тРНК. При взаимодействие комплекса (АК~АМФ)-АРС-аза с тРНК образуется аминоацил-тРНК, при этом выделяется свободный фермент и АМФ:
![]()
Похожие работы
... от иРНК, а с другой – обеспечивает образование небольшого количества более длинных полипептидов, могущих выполнять важную функциональную роль. Регуляция биосинтеза белков путем посттрансляционной модификации Посттрансляционная модификация белков менее распространена, чем процессинг РНК. Тем не менее известны случаи, когда при развитии некоторых вирусов трансляция полицистронной матрицы ...

... задачи были использованы методологии разработки программного обеспечения, детально рассмотренные в дипломном проекте, а также стандартные средства программных продуктов, представленных в настоящей работе. В связи с невозможностью использования прототипов специализированного шахматного программного обеспечения в ходе разработки были применены приемы экстремальной методологии разработки ПО. По ...
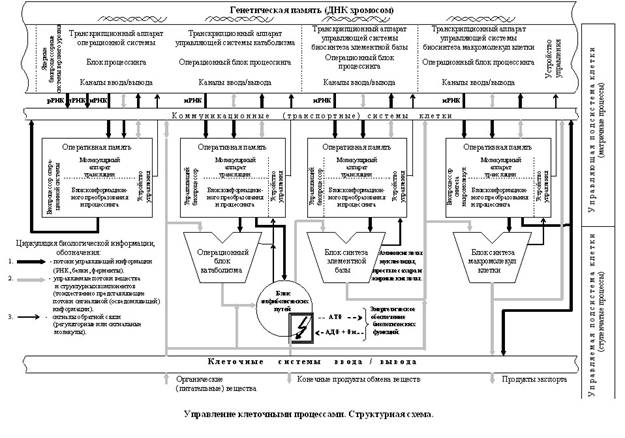
... её структурной и функциональной организации. Систематизируя отдельные информационные фрагменты и известные факты, в чем-то их переосмысливая, автор статьи сконструировал подобную схему и назвал её: “Информационное управление клеточными процессами. Структурная схема”. (Прилагается в конце статьи, с. 27). Структурная схема информационных процессов и сопряженных с ними процессов биохимических должна ...

... Министерство образования Российской Федерации Саратовский государственный технический университет Формульный компилятор методические указания к выполнению лабораторной работы по курсу «Теория вычислительных процессов и структур для студентов специальности ПВС Составил доцент кафедры ПВС Сайкин А.И. ...
0 комментариев