Навигация
4. Процессы трансляции
Второй этап матричного синтеза белка, собственно трансляцию, протекающую в рибосоме, условно делят на три стадии: инициацию, элонгацию и терминацию.
Инициация
В ходе инициации происходит образованиe комплекса, включающего Мет-тРНКiмет, мРНК и рибосому, где—тРНКiмет инициирующая метиониновая тРНК. В этом процессе участвуют не менее 10 факторов инициации, которые обозначают как elF (от англ. eukaryotic initiation factors) с указанием номера и буквы. У большинства мРНК-молекул эукариот 5¢-конец "кэпирован". Кэп представляет собой остаток метилгуанозилтрифосфата и, возможно, участвует в связывание РНК-молекул с 40S-субъединицей рибосомы. Первоначально 40S субъединица рибосомы соединяется с фактором инициации, который препятствует её связыванию с 60S субъединицей, но стимулирует объединение с тройным комплексом, включающим Мет-тРНКiмет, eIF-2 и ГТФ (Приложение 2 рис.1). Затем этот теперь уже более сложный комплекс связывается с 5'-концом мРНК при участии нескольких elF. Один из факторов инициации (elF-4F) узнаёт и присоединяется к участку "кэп" на молекуле мРНК, поэтому он получил название кэпсвязывающего белка. Прикрепившись к мРНК, 40S субъединица начинает скользить по некодирующей части мРНК до тех пор, пока не достигнет инициирующего кодона AUG кодирующей нуклеотидной последовательности. Скольжение 40S субъединицы по мРНК сопровождается гидролизом АТФ, энергия которого затрачивается на преодоление участков спирализации в нетранслируемой части мРНК. В эукариотических клеках некодирующие участки мРНК имеют разную длину, но обычно от 40 до 80 нуклеотидов, хотя встречаются области с протяжённостью более 700 нуклеотидов.
Достигнув начала кодирующей последовательности мРНК, 40S субъединица останавливается и связывается с другими факторами инициации, ускоряющими присоединение 60S субъединицы и образование 80S рибосомы за счёт гидролиза ГТФ до ГДФ и неорганического фосфата. При образовании полной рибосомы формируются два центра трансляции: донорный (пептидильный, P-центр) и акцепторный (аминоацильный, А-центр).
В Р-центре оказывается AUG-кодон мРНК с присоединённым к нему Мет-тРНКiмет, аминоацильный участок содержит аминоацил-тРНК, соединенную с соответствующим кодоном мРНК.
Элонгация
Самый продолжительный этап белкового синтеза — элонгация, в ходе которого рибосома с помощью аа-тРНК последовательно "читает" мРНК в виде триплетов нуклеотидов, следующих за инициирующим кодоном в направлении от 5' к З'-концу, наращивая полипептидную цепочку за счёт последовательного присоединения аминокислот. Присоединение соответствующей аминоацил-тРНК в А-участке требует точного узнавания кодона. Фактор элонгации EF1 образует комплекс с ГТФ и молекулой аминоацил-тРНК. Благодаря этому аминоацил-тРНК может присоединиться к рибосоме. При этом произойдет высвобождение комплекса EF1-ГДФ и фосфата (приложение 2, рис. 2). Комплекс EF1-ГДФ затем вновь превращается в EF1-ГТФ при участии других свободных белковых факторов и ГТФ.
a-Аминогруппа новой амино-ацил-тРНК в участке А осуществляет нуклеофильную атаку этерефицированной карбоксильной группы пептидил-тРНК, занимающей P-участок. Эта реакция катализируется пептидилтрансферазой – белковым компонентом, входящим в состав 60S-рибосомной субъединицы.
После удаления пептидильного остатка с тРНК в Р-участке свободная молекула тРНК быстро покидает P-участок. Комплекс ГТФ с EF2 участвует в процессе транслокации новообраованной пептидил-тРНК из А-участка в Р-участок. При этом происходит гидролиз ГТФ, используемого в качестве кофактора EF2, до ГДФ и фосфата (рис.3). В результате транслокации вновь сформированная пептидил-тРНК и соответствующий ей кодон переходят в Р-участок, освобождая А-участок для нового цикла узнавания следующего кодона соответствующей молекулой аминоацил-тРНК и элонгации.
Терминация
Терминация представляет собой завершение синтеза полипептидной цепи и освобождение ее от рибосомы (рис. 4). После многих циклов элонгации, в результате которых синтезируется полипептидная цепь белка, в А-участоке появляется терминирующий или нонсенс-кодон. В норме отсутствуют молекулы тРНК, способные узнавать нонсенс-кодоны. Появление в А-участке терминирующего кодона распознается так называемыми факторами высвобождения (R-факторами). RА при участии ГТФ и пептидилтрансферазы обеспечивают гидролиз связи между полипептидом и молекулой тРНК, занимающей P-участок. После гидролиза и высвобождения синтезируемого полипептида и тРНК 80S-рибосома диссоциирует на 40S- и 60S-субъединицы.
Одну и туже цепь мРНК могут транслировать одновременно множество рибосом. Рибосомы, расположенные на одной молекуле мРНК, образуют полисому.
5. Регуляция биосинтеза белка на этапе трансляции
Лимитирующей стадией процесса трансляции является ее инициация. Наиболее подробно описан процесс изменения скорости инициации трансляции в результате фосфорилирования фактора инициации IF2. Реакция катализируется ферментом IF2-киназой, причем присоединение фосфатной группы инактивирует фактор инициации. Этот феномен был изучен на примере синтеза гемоглобина в ретикулоцитах. Оказалось, что активация IF2-киназы происходит за счет ее фосфорилирования цАМФ-зависимой протеинкиназой. Взаимодействие этой протеинкиназы с цАМФ и ее активацию блокирует гем, выполняя тем самым негативный контроль синтеза гемоглобина.
К лекарственным веществам, эффективно влияющим на синтез белка, относятся антибиотики. Большинство антибиотиков противобактериального действия ингибируют процессы трансляции. Такие антибиотики, как норвалин и индомицин, препятствуют образованию аминоацил-тРНК; стрептомицин, неомицин, конвалин, ауринтрикарбоновая кислота ингибируют инициацию трансляции; тетрациклин и стрептомицин ингибируют элонгацию, препятствуя связыванию аминоацил-тРНК с А-центром рибосомы. Пептидилтрансферазная реакция блокируется пуромицином и хлорамфениколом, а транслокация – эритромицином и виомицином.
Заключение
Трансляция крайне важный процесс, и нарушение или выпадение любого звена, участвующего в синтезе белка, почти всегда приводит к развитию патологии, причем клинические проявления болезни будут определяться природой и функцией белка, синтез которого оказывается нарушенным (структурный или функциональный белок). Иногда синтезируются так называемые аномальные белки как результат действия мутагенных факторов и, соответственно, изменения генетического кода (например, гемоглобин при серповидно-клеточной анемии). Последствия этих нарушений могут выражаться в развитии самых разнообразных синдромов или заканчиваться летально. Следует отметить, что организм располагает мощными механизмами защиты: подобные изменения генетического аппарата быстро распознаются специфическими ферментами — рестриктазами, измененные последовательности вырезаются и вновь замещаются соответствующими нуклеотидами при участии полимераз и лигаз.
Список литературы
1. Биохимия человека в 2 томах, Т 2//Р. Марри 2009 г.
2. Биохимия// Комов В.П., 2004г.
3. Биологическая химия//Шапиро Я.С., 2004г.
4. Биохимия//Северин Е.С., 2005г.
5. Биологическая химия//Березов Т.Т., Коровкин Б.Ф., 1990 г.
Приложение 1
Таблица 1. Вырожденность кода аминокислот
| Аминокислота | Число кодонов | Аминокислота | Число кодонов |
| Мет Трп | 1 | Иле | 3 |
| Асн Асп Цис Глн Глу Гис Лиз Тир Фен | 2 | Ала Гли Про Тре Вал | 4 |
| Арг Сер Лей | 6 |
Таблица 2. Генетический код
| Первое основание | Второе основание | Третье основание | |||
| U | U Фен Фен Лей Лей | C Сер Сер Сер Сер | A Тир Тир * * | G Цис Цис * Три | U С A G |
| C | Лей Лей Лей Лей | Про Про Про Про | Гис Гис Глн Глн | Арг Арг Арг Арг | U С A G |
| A | Иле Иле Мет Мет | Тре Тре Тре Тре | Асн Асн Лиз Лиз | Сер Сер Арг Арг | U С A G |
| G | Вал Вал Вал Вал | Ала Ала Ала Ала | Асп Асп Глу Глу | Гли Гли Гли Гли | U С A G |
Приложение 2
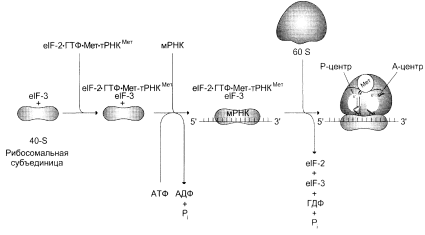
Рисунок 1
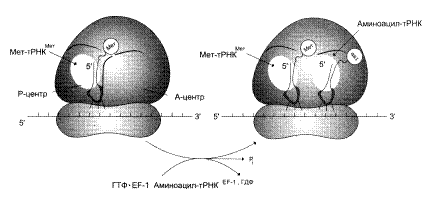
Рисунок 2
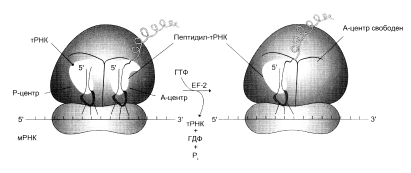
Рисунок 3
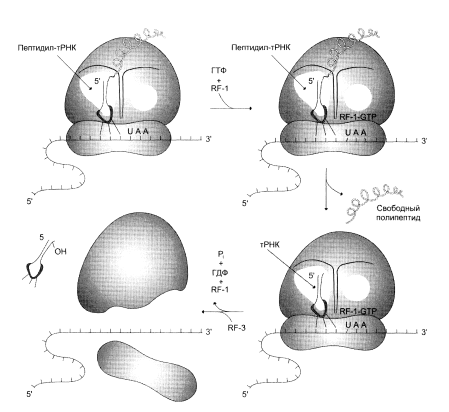
Рисунок 4
Похожие работы
... от иРНК, а с другой – обеспечивает образование небольшого количества более длинных полипептидов, могущих выполнять важную функциональную роль. Регуляция биосинтеза белков путем посттрансляционной модификации Посттрансляционная модификация белков менее распространена, чем процессинг РНК. Тем не менее известны случаи, когда при развитии некоторых вирусов трансляция полицистронной матрицы ...

... задачи были использованы методологии разработки программного обеспечения, детально рассмотренные в дипломном проекте, а также стандартные средства программных продуктов, представленных в настоящей работе. В связи с невозможностью использования прототипов специализированного шахматного программного обеспечения в ходе разработки были применены приемы экстремальной методологии разработки ПО. По ...
... её структурной и функциональной организации. Систематизируя отдельные информационные фрагменты и известные факты, в чем-то их переосмысливая, автор статьи сконструировал подобную схему и назвал её: “Информационное управление клеточными процессами. Структурная схема”. (Прилагается в конце статьи, с. 27). Структурная схема информационных процессов и сопряженных с ними процессов биохимических должна ...

... Министерство образования Российской Федерации Саратовский государственный технический университет Формульный компилятор методические указания к выполнению лабораторной работы по курсу «Теория вычислительных процессов и структур для студентов специальности ПВС Составил доцент кафедры ПВС Сайкин А.И. ...
0 комментариев