Навигация
Ионные механизмы потенциала покоя
15072
знака
0
таблиц
2
изображения
Ионные механизмы потенциала покоя
Содержание
Введение
Идеальная клетка
Ионное равновесие
Электрическая нейтральность
Влияние внеклеточного калия и хлора на мембранный потенциал
Литература
Введение
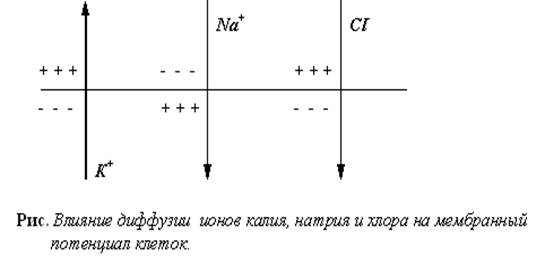
В состоянии покоя на мембране нейронов поддерживается постоянный электрический потенциал, при этом внутриклеточная среда заряжена отрицательно по отношению к внеклеточной. Поскольку внутри клетки больше ионов калия и меньше ионов натрия и хлора, чем снаружи, то каждый из этих ионов стремится проникнуть сквозь мембрану в направлении своего электрохимического градиента, то есть ионы калия стремятся наружу, а ионы натрия и хлора - внутрь клетки. Этому перемещению препятствует мембранный потенциал. Если представить себе упрощенную модель клетки, мембрана которой проницаема лишь для калия и хлора, то в ней возможен такой баланс между концентрационными градиентами и мембранным потенциалом, при котором суммарный ток каждого из этих ионов равен нулю. Тогда мембранный потенциал будет в точности равен равновесному потенциалу для этих двух ионов.
Если в такой модели изменить внеклеточную концентрацию калия, то изменится равновесный потенциал для калия, а следовательно и мембранный потенциал. Изменение внеклеточной концентрации хлора, напротив, приведет к соответствующему изменению его внутриклеточного уровня. При этом ни равновесный потенциал для хлора, ни мембранный потенциал не изменятся.
Мембрана реальной клетки проницаема также для натрия. В состоянии покоя ионы натрия непрерывно перемещаются внутрь клетки, снижая таким образом отрицательный заряд на мембране. Поскольку при этом калий уже не находится в равновесии, он начинает вытекать из клетки. Если бы не механизмы компенсации, то такое перемещение натрия и калия привело бы к изменению их внутриклеточных концентраций. Этого не происходит благодаря работе натрий-калиевого насоса-обменника, переносящего ионы натрия наружу и ионы калия внутрь клетки в пропорции 3:2. Мембранный потенциал покоя зависит от калиевого и натриевого равновесного потенциала, относительной проницаемости клеточной мембраны для этих ионов, а также от стехиометрии натрий-калиевого насоса. В состоянии покоя скорость переноса натрия и калия насосом в точности компенсирует пассивную утечку этих ионов. Натрий-калиевый обменник вносит также непосредственный вклад в мембранный потенциал (в размере нескольких милливольт), поскольку он перемещает больше положительных зарядов наружу, чем внутрь клетки.
В зависимости от механизмов транспорта, равновесный потенциал для хлора может быть как более, так и менее отрицательным по отношению к потенциалу покоя. Наличие значительной хлорной проницаемости играет важную роль в обеспечении электрической стабильности многих клеток, несмотря на то, что распределение ионов хлора не столь важно для формирования потенциала покоя.
Возникновение электрических сигналов в нервной клетке происходит в основном за счет изменения проницаемости мембраны для таких ионов, как натрий и калий. Увеличение проницаемости мембраны позволяет ионам перемещаться внутрь или наружу клетки в направлении своих электрохимических градиентов. Изменения проницаемости зависят от активации ионных каналов. Перемещение ионов через каналы в мембране приводит к изменению заряда на ней и, следовательно, мембранного потенциала. Чтобы понять принцип возникновения электрического сигнала, необходимо уяснить сущность ионных градиентов на мембране, а также то, как они влияют на потенциал покоя.
Идеальная клетка
Начнем с рассмотрения модели идеальной клетки, показанной на рис.1.1. В этой клетке содержатся ионы калия, натрия и хлора, а также крупные анионы. Клетка помещена в раствор, содержащий ионы натрия и калия. Другие ионы, присутствующие в реальных клетках, не играют важной роли в формировании потенциала покоя и потому не рассматриваются в данной модели. Концентрации ионов внутри и снаружи клетки приблизительно соответствуют средним величинам, полученным на клетках лягушки. В клетках птиц и млекопитающих эти значения несколько выше, а у таких морских беспозвоночных, как кальмар - значительно выше, чем у лягушки (см. табл. 1.1). В настоящей модели рассматривается клеточная мембрана, проницаемая для калия и хлора, но не для натрия и внутриклеточных анионов. Для того чтобы клетка пребывала в стабильном состоянии, необходимо выполнение трех условий:
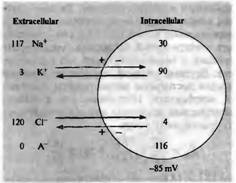
Рис.1.1 Распределение ионов в идеальной клетке.
Мембрана проницаема для К и Cl, но непроницаема для Na и внутриклеточных анионов (А). Концентрационный градиент ионов калия способствует выходу этих ионов из клетки (черная стрелка); градиент потенциала стремится перенести ионы калия внутрь клетки (серая стрелка). В состоянии покоя эти две силы уравновешивают друг друга. Концентрационный и электрический градиенты для хлора имеют противоположные направления. Ионные концентрации выражены в миллимолях (ммоль).
1. Как внутриклеточная, так и внеклеточная среды должны быть электрически нейтральными. Так, например, раствор не может состоять из одних лишь ионов хлора. Отрицательный заряд этих ионов должен компенсироваться такими положительными ионами, как натрий или калий (иначе взаимное отталкивание одинаково заряженных частиц приведет к разрушению раствора).
2. Клетка должна находиться в осмотическом равновесии. В противном случае вода будет проникать в клетку (или вытекать из нее) до тех пор, пока равновесие не будет достигнуто. Осмотическое равновесие наступает при условии равенства суммарной концентрации растворенных частиц внутри и вне клетки.
Похожие работы
... бы гораздо медленнее, чем в действительности. Причина этого в том, что общая проводимость мембраны в покое достаточно мала, и выход накопленных в результате потенциала действия положительных ионов из клетки занял бы несколько миллисекунд, а то и десятки миллисекунд. В действительности, возврат к уровню потенциала покоя происходит гораздо быстрее благодаря значительному увеличению другой мембранной ...
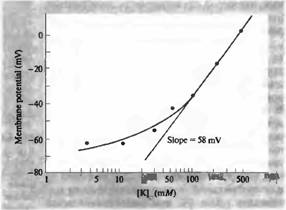
... мембранной проницаемости для натрия, экспериментальные данные расходятся с прямой линий, особенно при низких концентрациях калия. Из уравнения постоянного поля следует один важный принцип. Мембранный потенциал зависит от проводимости (или проницаемости) мембраны для важнейших ионов, а также от их равновесных потенциалов. В реальных клетках проводимости для калия и хлора сравнительно· высоки, ...
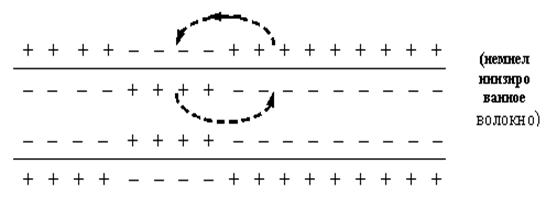
... вдоль волокна без затухания (бездекрементное проведение). Это обусловлено тем, что локальные токи только деполяризуют мембрану до критического уровня, а потенциалы действия в каждом участке мембраны поддерживаются независимыми ионными потоками, перпендикулярными к направлению распространения возбуждения. Скорость уменьшения мембранного потенциала до критического уровня зависит от разности ...
е реобазы и хронаксии ввёл в физиологию Л. Лапик в 1909, определяя зависимость между силой тока и длительностью его действия при изучении наименьшего (порогового) эффекта возбудимых тканей. Реобаза, как и хронаксия, даёт представление о возбудимости тканей и органов по порогу силы и длительности действия раздражения. Реобаза соответствует порогу раздражения и выражается в вольтах или миллиамперах. ...
0 комментариев