Навигация
Картирование сложно наследуемых признаков человека
41913
знаков
0
таблиц
6
изображений
Картирование сложно наследуемых признаков человека
Введение
За последние несколько лет генетика человека достигла значительных успехов. Конечно, еще с момента переоткрытия законов Менделя было очевидно, что человек подчиняется тем же законам наследственности, что и другие организмы. Более того, именно анализ наследования семейных заболеваний убедительно свидетельствовал в пользу менделизма. Однако, несмотря на это многообещающее начало, генетика человека намного отстала от частной генетики таких организмов, как дрозофила и кукуруза, по двум главным причинам. Во-первых, люди не лабораторные животные, с которыми можно проводить скрещивания, адекватные целям эксперимента, и, во-вторых, обнаружено лишь небольшое число генетических маркеров, степень гетерозиготности по которым, позволяла рассматривать встречающиеся в естественной популяции браки как информативные для анализа сцепления.
Разработка метода рекомбинантных ДНК явилась импульсом для развития генетики человека. Было высказано предположение, что генетический полиморфизм на уровне последовательностей ДНК, который можно легко наблюдать на примере полиморфизма длины рестрикционных фрагментов, – достаточно частое явление и соответствующие аллельные варианты могут использоваться как генетические маркеры, позволяющие проводить систематическое изучение наследственности человека, включая построение полной картины его генома.
На сегодняшний день известно более 2500 ПДРФ-маркеров. Анализ некоторых из них в родословных с больными выявил ПДРФ-маркеры, проявляющие тесное генетическое сцепление с мышечной дистрофией Дюшенна, болезнью Гентингтона, муковисцидозом 7–10, поликистозом почек у взрослых, ретинобластомой 12, ранней семейной формой болезни Альцгеймера, биполярным аффективным психозом в большой родословной амишей, нейрофиброматозом Реклингаузена, множественной эндокринной неоплазией типа 2А и семейным полипозом. Более того, значительный прогресс в построении генетической карты человека обещает сделать процесс поиска сцеплений более эффективным за счет замены случайного выбора ПДРФ систематическим анализом всех ПДРФ, диспергированных по геному человека.
Достигнутые успехи убеждают нас в том, что для любого заболевания человека может быть найден сцепленный с ним ПДРФ-маркер при условии, что это заболевание:
1) наследуется как простой менделеевский признак – рецессивный или доминантный;
2) возникает вследствие мутации в уникальном генном локусе;
3) достаточно распространено, чтобы можно было собрать необходимое количество семей с несколькими пораженными.
К сожалению, очень многие из интересующих нас наследственных признаков человека обусловлены генетическими причинами, менее подходящими для такого анализа. Затруднения заключаются в следующем:
1) неполная пенетрантность, вследствие которой не все индивиды – носители мутантного генотипа, проявляют фенотип, отличный от нормы;
2) фенокопии, вследствие которых индивиды с нормальным генотипом могут проявить мутантный фенотип по причинам негенетического характера;
3) генетическая гетерогенность, благодаря которой мутации по нескольким различным генетическим локусам могут дать вполне сходную клиническую картину;
4) генные взаимодействия, вследствие которых фенотип является результатом функционирования аллелей нескольких локусов;
5) редкость заболевания, которая может затруднить или сделать невозможным анализ семей с несколькими больными индивидами.
Эти проблемы обычны при анализе таких хорошо изученных объектов, как бактерии, дрожжи, нематоды и плодовые мушки. По-видимому, и человек в этом отношении не является исключением. Чтобы преодолеть указанные трудности, генетики обычно работают с чистыми линиями, несущими мутацию по единичному локусу, культивируя огромное количество особей в контролируемых условиях и проводя целенаправленные скрещивания. В генетике человека приходится, однако, анализировать уже имеющиеся браки.
В этой работе мы даем обзор основных принципов, на основе которых планируется изучение генетического сцепления у человека в случаях, когда возникают указанные осложнения. Акцент будет сделан на практических вопросах отбора семей с целью максимизации вероятности обнаружения сцепления. Мы не будем подробно останавливаться на вычислительных и чисто алгебраических аспектах такого анализа. Необходимо заметить, что оптимальное планирование анализа сцепления существенно зависит от точности модели наследования заболевания, а это не часто бывает известно заранее. Если генетика в действительности очень сложна, анализ сцепления явно будет неуспешным. Поэтому выбор плана зависит в определенной степени от знания возможных осложнений. Заметим, что точное планирование значительно увеличивает мощь анализа сцепления при изучении заболеваний со сложным типом наследования.
1. Основы анализа сцепления
Основы анализа генетического сцепления достаточно полно изложены в работе Отта. Мы лишь суммируем здесь основные понятия анализа правдоподобия.
Предположим, что мы хотим сравнить рабочую гипотезу Н с альтернативной «нулевой» гипотезой Но. Гипотеза Н, к примеру, может утверждать, что локус ПДРФ сцеплен с заболеванием с 10%-ной частотой рекомбинаций, в то же время по гипотезе. Но сцепление может вовсе отрицаться. После сбора семейных данных мы подсчитываем две вероятности: Р – вероятность того, что наблюдаемые данные не противоречат основной гипотезе Н, когда она истинная, и Ро-вероятность того, что те же данные могли иметь место, когда верна гипотеза Но. По отношению Р / Ро можно судить о том, насколько более вероятно появление наблюдаемых данных при условии истинности Н, чем при условии истинности Но – Если это отношение превосходит какой-нибудь достаточно большой порог Т, мы отвергаем Но в пользу гипотезы Н.
По соглашению, в генетике человека предпочитают использовать десятичные логарифмы отношений вероятностей, именуемые лод-баллами. Отношения вероятностей для независимых выборок могут быть объединены путем перемножения, и, таким образом, лод-баллы, полученные по независимым выборкам, могут просто суммироваться.
При планировании анализа возникают два вопроса.
1. Порог: какой должна быть величина порога Т, чтобы снизить до приемлемой величины ошибочно отвергнуть нулевую гипотезу Н0, в случае, когда она в действительности верна?
2. Необходимая величина выборки: как велика должна быть выборка для того, чтобы можно было с большой достоверностью принять гипотезу Н в случае, когда она действительно верна?
В генетике человека принятая величина порога для единичного наблюдения сцепления соответствует отношению шансов не меньше чем 1000:1 и значению лод-балла 3,0. Этот порог не такой строгий, как может показаться. Два случайным образом выбранных локуса в геноме человека, вероятнее всего, располагаются в разных хромосомах или в разных хромосомных плечах, если речь идет об одной хромосоме. При грубой оценке имеется шансов 50:1 против сцепления. Даже если, согласно наблюдаемым данным, вероятность сцепления в 1000 раз превышает вероятность его отсутствия, тот факт, что в общем случае отсутствие сцепления в 50 раз более правдоподобно, чем его наличие, означает, что лод-балл, равный 3, соответствует истинной величине отношения шансов за сцепление только 20:1. Другими словами, при значении лод-балла, равном 3, в одном из 20 случаев может быть ложный результат. При величине лод-балла, равной 4, результат за наличие сцепления будет фиктивным в одном случае из 200.
Необходимый размер выборки может быть установлен подсчетом ожидаемого вклада каждой семьи в общую сумму лод-баллов. При заданной структуре семьи подсчитывается «ожидаемое значение лод-балла», или ELOD. Необходимо набрать достаточное количество семей, чтобы сумма ожидаемых лод-баллов значительно превысила порог, требуемый для принятия гипотезы о наличии сцепления.
Сила метода правдоподобия – в его гибкости. При наличии генных взаимодействий, к примеру, можно проверить гипотезу, согласно которой для проявления болезни необходимо наличие мутаций одновременно по обоим локусам. В случае же с генетической гетерогенностью можно проверить гипотезу о том, что для проявления заболевания достаточно мутации по любому из двух локусов. В данной ситуации это означает, что в каждой семье будет наблюдаться сцепление только с одним из двух локусов.
Похожие работы
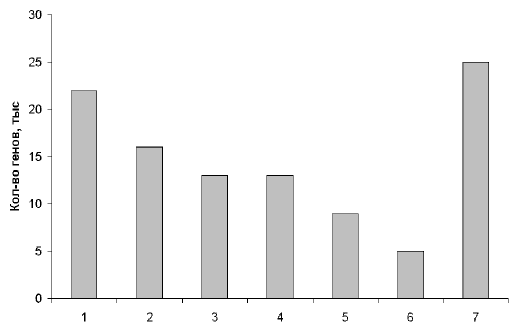
... , в нашем геноме есть масса повторяющихся участков. Действительно, очень интересно, почему человек - "венец эволюции" имеет огромную долю "неработающего" генома. Рис. 2. Примерное распределение генов человека по их функциям. 1 – производство клеточных материалов; 2 – производство энергии и ее использование; 3 – коммуникации внутри и вне клеток; 4 – защита клеток от инфекций и повреждений; 5 – ...




... только те страны и компании, которые в состоянии выложить за это значительные деньги. Бедные же страны, например африканские, могут оказаться отрезанными от достижений прогресса в геномике.Изучая проект «Геном человека» учёныё столкнулись с величайшим открытием века. Коротко об этом: Электромагнитный апокалипсис. Как это ни странно звучит, но в последние десятилетия в науке открылись такие ранее ...
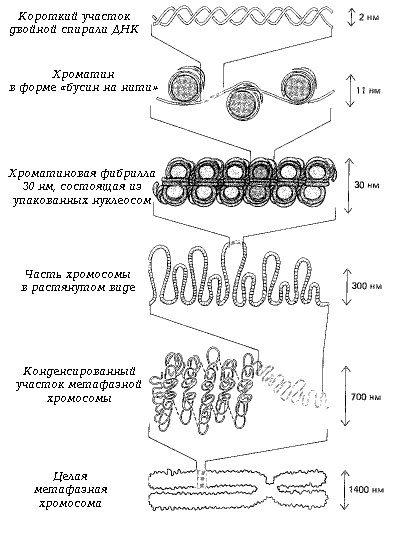
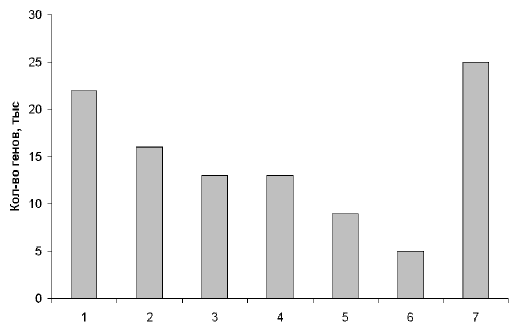
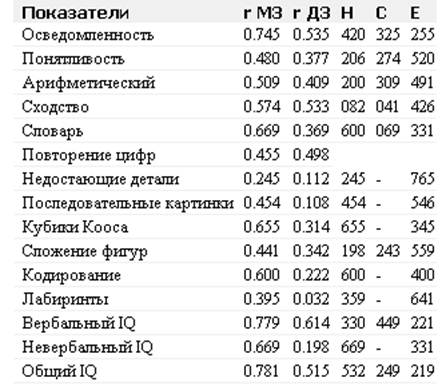
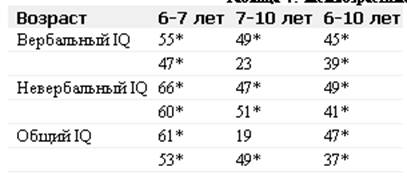
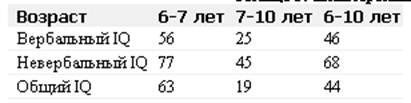
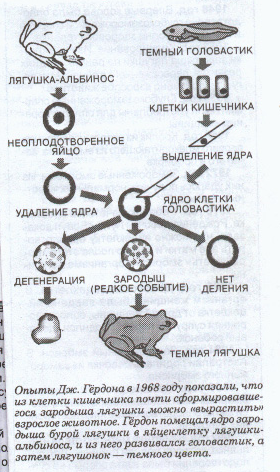
... отклонениям в развитии организма. Например, цитогеническим методом установлены причины болезни Дауна у человека. Эту болезнь вызывает наличие лишней хромосомы в одной из хромосомных пар. В генетике человека нередко применяется генеалогический метод - изучение родословных. Он позволяет выявить доминированность или рецессивность гена, сцепленность генов между собой и с полом. В клетках человека ...
... придется смириться с определенным числом химически индуцированных мутаций, поскольку общество не может отказаться от тех преимуществ, которые дают ему достижения современной химии. В будущем человечеству придется столкнуться с увеличением частоты спонтанных мутаций. Оно приведет к соответствующему увеличению численных и структурных хромосомных аберраций и наследственных заболеваний, ...
0 комментариев