Навигация
СОКРАТИТЕЛЬНЫЕ И ЦИТОСКЕЛЕТНЫЕ БЕЛКИ НЕРВНОЙ ТКАНИ
53771
знак
0
таблиц
1
изображение
3. СОКРАТИТЕЛЬНЫЕ И ЦИТОСКЕЛЕТНЫЕ БЕЛКИ НЕРВНОЙ ТКАНИ
Рассматривая сократительные и цитоскелетные белки, входящие в состав нейрональных и нейроглиальных клеток, следует отметить, что не все они отличны от сократительных белков других тканей.
Пока нет достаточных оснований, например, считать нейроспецифическими миозин и актин нервной ткани.
Специфические сократительные белки обеспечивают динамичность вообще и механическую подвижность нервной ткани, участвуя в самосборке и распаде специфических структур – микротрубочек, нейрофиламентов и других пре-синаптических и постсинаптических образований, в переносе различных соединений между разными областями нейрона, а также в поддержании и модуляции пространственного положения частей нейрона.
Микротрубочки и нейрофиламенты являются важнейшими структурными образованиями нервных клеток, обладающими как скелетными, так и сократительными свойствами. Они принимают непосредственное участие в прямом и ретроградном транспорте клеточных органелл, нуклеиновых кислот, белков, сложных липидов, липо- и гликопротеинов и их предшественников, а также ряда других метаболитов по аксону от тела нейрона до синаптических окончаний. Кроме того, они участвуют в движении метаболитов в различных субклеточных структурах тела нейронов. Они претерпевают изменения при различных функциональных состояниях и, являясь динамическими структурами, могут влиять на топографию поверхности нейрона и на мозаичность нейрональных мембран. Не менее важно и их значение в самосборке микроструктур и надмолекулярных комплексов, а также в поддержании определенной конфигурации микроструктур.
Микротрубочки представляют собой образования цилиндрической формы, диаметр которых достигает 24 нм, а наибольшая длина соизмерима с длиной отростков нейронов. Основная масса белка, входящего в состав микротрубочек, приходится на долю нейротубулина. Его количество достигает 15% от суммы растворимых белков мозга. В период формирования и увеличения размеров мозга содержание в нем нейротубулина еще выше.
Нейротубулин является димером, в его состав входят 2 субъединицы – а-тубулин и р-тубулин. Молекулярная масса мономеров тубулина составляет соответственно 53 и 57 кД.
Нейротубулин является кислым белком, в его составе содержится около 20% глутаминовой и аспарашновой кислот.
В микротрубочках нейротубулин находится в виде спиральных полимеров, состоящих из 10–14 молекул нейротубулина. Формирование полимерной трубчатой структуры протекает с потреблением макроэргов – за счет ГТФ. Сам нейротубулин обладает ГТФазной активностью. В полимеризации тубулина принимает также участие специальный белок сборки тубулина – Т-фактор. Сборка и разборка микротрубочек in vivo происходит очень быстро. Подавляется сборка микротрубочек известными ядами – колхицином, винбластином и винкрестином.
Нейротубулин даже при высокой степени очистки обладает фосфокиназной и протеинкиназной активностью, так как эти ферменты, по-видимому, являются обязательными компонентами нейротубулина.
Сравнение пептидных карт тубулинов, выделенных, например из микротрубочек мозга куриного эмбриона и сперматозоидов морского ежа, показало значительную степень их иденточности. Это дает основание считать тубулины относительно стабильными белками в эволюционном плане.
Нейрофиламенты образуются из спирально скрученных нитевидных образований, диаметр их колеблется в пределах 5–12 нм. Филаменты размером 5–6 нм называются микрофиламентами. Нейрофиламенты содержат сравнительно много актомио-зинподобных белков, особенно актина в нитевидной, F‑форме. Образование актиновых нитей в нейрофиламентах состоит в полимеризации молекул глобулярного G‑актина с Мг 46 кД, протекает за счет энергии гидролиза АТФ до АДФ и подавляется алколоидом – цитохалазином В. В состав нейрофиламентов входит также коллаген, который может быть удален с помощью коллагеназы. Обнаружена видовая специфичность белков нейрофиламентов в мозге человека по сравнению с мозгом животных.
Актомиозинподобные белки можно отнести к сократительным белкам нервной ткани. Они были извлечены различными способами из целого мозга и отдельных частей нервной системы. В развивающемся мозге и в культуре нейронов содержание актиноподобных белков достигает 8%, а миозинподобных белков – 0,5%. У взрослых животных количество последних несколько меньше. По аминокислотному составу и первичной структуре актиноподобный белок близок к актину из мышц.
Актомиозинподобные белки участвуют в аксональном токе и освобождении трансмиттеров в синапсах. Кроме того, они были обнаружены в конусе роста, где их содержание довольно высоко в отличие от низкого содержания этих белков в культуре нейробластов. Имеются косвенные данные о том, что в нативном состоянии часть актиноподобных белков находится в комплексе с миозинподобными белками, причем эти комплексы чувствительны к митогенетическим ядам, в то время как в отдельности указанные белки малочувствительны к этим агентам.
К актомиозинподобным белкам ЦНС относится нейростенин. Он состоит из двух белков – нейрина и стенина. Взаимодействуя между собой, они образуют комплекс – нейростенин с Мг – 47–50 кД. Он имеет много общего с актомиозином мышцы по структуре и по функциям, хотя и не идентичен ему.
Нейростенин обладает АТФазной активностью и активируется ионами Са+ и Mg+. Количество нейростенина составляет около 1–1,5% от общего белка мозга; однако в синаптических образованиях его содержание достигает 8–10%. Нейрин локализован преимущественно в пресинаптических мембранах, а стенин – на наружной поверхности мембран везикул. С формированием нейростенина в присутствии АТФ и ионов Са+ связывают предположительно контакт везикул с пресинаптическими мембранами. Полагают, что сократительные белки мозга, в том числе нейростенин, участвуют в раскрытии везикул и выходе нейромедиатора в цитоплазму и синаптическую щель. В «плавлении» мембраны везикул, происходящем при выбросе медиатора, важную роль играют также синапсины и другие Са-связывающие белки, описанные выше.
Большой интерес представляет другой сократительный белок нейронов – кинезин. Этот недавно открытый цитоплазматический транслокатор является «механохимической» АТФа-зой, способной обеспечивать скольжение внутриклеточных органелл вдоль микротрубочек. Он служат одним из двигателей антероградного аксонального тока.
Кинезин из мозга крупного рогатого скота состоит из двух полипептидных цепей, неодинаковых по первичной структуре. Он образует прочный комплекс, в котором субъединицы связаны нековалентно. Молекула нативного кинезина состоит, по-видимому, из двух пар указанных субъединиц с Мг = 384 кД. В клетке обе субъединицы связаны с микротрубочками. Чистый кинезин обладает слабой АТФазной активностью, которая, однако, возрастает в несколько раз в присутствии микротрубочек. Молекулы АТФ соединяются с тяжелой субъединицей кинезина, которая является каталитически активной. Энергии, высвобождающейся при гидролизе АТФ кинезином, достаточно для обеспечения передвижения даже крупных внутриклеточный органелл в аксонах. В последнее время описан белок, подобный кинезину, но неидентичный ему, обеспечивающий ретроградный аксональный ток, – динеин.
Другой белок – спектрин, обнаруженный сначала в мембранах эритроцитов, где он составляет до 30% всех мембранных белков, а затем и в клетках многих органов и тканей, представляет собой компонент цитоскелета клетки. Длинная фибриллярная молекула спектрина состоит из двух полипептидных цепей. Такие молекулы образуют субмембранную сеть филаментов на внутренней поверхности цитоплазматической мембраны, которая через молекулы другого белка – анкирина взаимодействуют с другими белками цитоскелета, что снижает подвижность белков в плоскости мембраны. В ткани головного мозга спектрин участвует также в распределении ионов К+ и Na+ по поверхности мембран возбудимых клеток.
Белок клатрин впервые был выделен из мембран так называемых окаймленных пузырьков, принимающих участие в эндоцитозе и быстром внутриклеточном транспорте веществ. Этот фибриллярный белок; ау – гибридная форма, обозначавмая как белок 14–3–1, и уу – нейроспецифический изоэнзим енолазы, локализованный только в нейронах.
Молекулярная масса белка 14–3–2 близка к 80 кД. Как и белок S‑100, он содержит относительно много дикарбоновых кислот. Интересно, что эта изоформа термостабильна до температуры 50°С, Значительно различаются и периоды полужизни изоферментов енолазы: для уу-димера он равен 320 мин, а для аа-димера – 15 мин.
Белок 14–3–2 широко распространен в ЦНС и ПНС млекопитающих и птиц. Его количество составляет около 1,5% от общих растворимых белков мозга. В отличие от белка S‑100 он локализован главным образом в нейронах, а в клетках нейрог-лии его содержание незначительно.
Белок 14–3–2 сосредоточен в сером веществе больших полушарий. В других органах и тканях человека этот белок отсутствует или содержится в количествах, в 50–100 раз меньших. Иммуно-химическим методом показано, что в постнатальный период развития головного мозга крыс белок 14–3–2 наиболее интенсивно синтезируется в гиппокампе и синаптических мембранах. В опытах с дегенерацией зрительного нерва было обнаружено снижение содержания и интенсивности метаболизма белка 14–3–2.
По своей внутриклеточной локализации белок 14–3–2 является в основном цитоплазматическим и присутствует в цитоплазме как нейронов, так и периферических нервов. Он транспортируется с помощью медленного аксотока. Обнаружено три типа нейронов: а) приобретающие белок 14–3–2 раньше других в ходе онтогенеза и сохраняющие его постоянно; б) имеющие белок 14–3–2 только в определенный период онтогенеза; в) не имеющие этого белка.
Открытие нейроспецифического белка 14–3–2 явилось в свое время важным событием, поскольку, во-первых, был обнаружен новый специфический нейрональный белок, во-вторых, показано, что нейроспецифические белки могут представлять собой мозговые изоферменты уже известных энзимов.
В настоящее время идентифицирован целый ряд нейроспецифических изоферментов; среди них можно назвать мозговые формы альдолазы, арилсульфатазы, ВВ-изо-зим креатинкиназы и многие другие.
Похожие работы
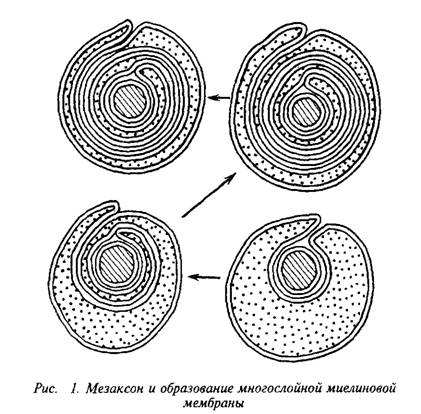
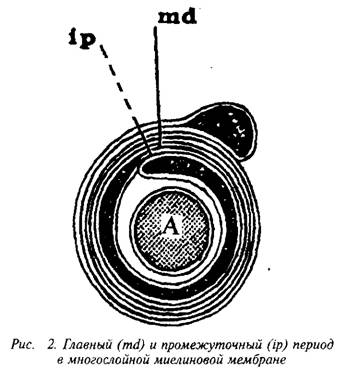
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
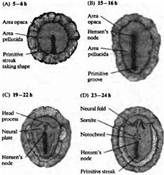
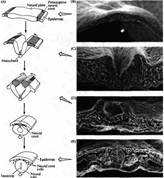
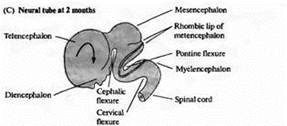
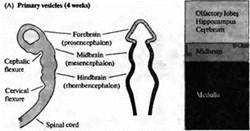
... уровне направленные мутации, что открывает дорогу к обнаружению важных генов позвоночных, которые могут иметь гомологов у некоторых видов более просто устроенных беспозвоночных. Развитие нервной системы в раннем периоде В раннем периоде морфогенеза позвоночных область гаструлы, из которой в дальнейшем формируется нервная система, представляет собой простую полоску эктодермы (рис. 1). ...
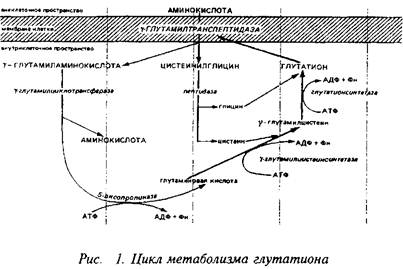
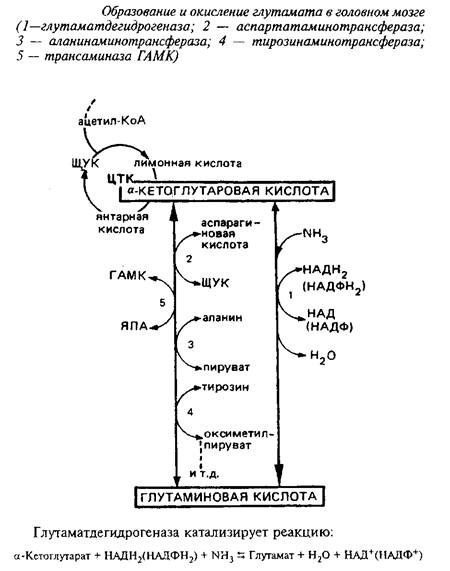
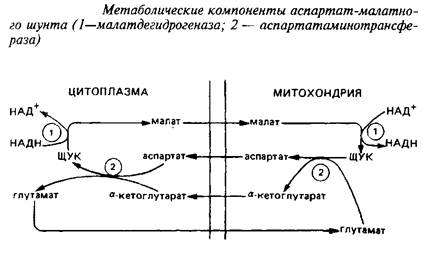
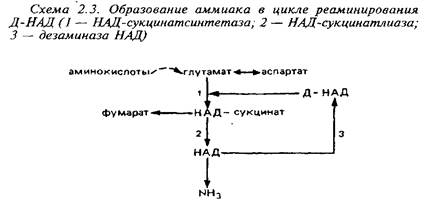
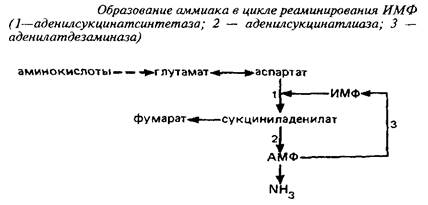
... видов животных. Региональные различия в концентрации метионина невелики. Влияние диеты на концентрацию метионина в мозге также незначительно из-за конкурентных отношений с нейтральными аминокислотами за транспортные системы. Метионин в пуле свободных аминокислот утилизируется на 80% для синтеза белка. Метаболизм свободного метионина до цистеина начинается с образования S-аденозилметионина, ...
... в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой. Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение ...
0 комментариев