Навигация
Встраивание белков в мембрану
36107
знаков
0
таблиц
4
изображения
2. Встраивание белков в мембрану
2.1 Сигнальная гипотеза
Белки встраиваются в мембрану разными способами, но детали этого процесса во многих случаях еще не установлены. Для объяснения механизма встраивания предложены две модели: сигнальная гипотеза и мембранная триггерная гипотеза. В сигнальной гипотезе предполагается, что белок включается в мембрану параллельно его трансляции на мРНК в полирибосомах; это так называемое котрансляционное включение. Когда лидерная последовательность выходит из рибосомы, она выявляется некой сигнал-распознающей частицей (СРЧ), которая блокирует дальнейшую трансляцию на уровне примерно 70 аминокислот, 40 из которых остаются в большом рибосомном комплексе, а 30 экспонированы в среду. СРЧ содержит шесть белков, с ней ассоциирована 7S-РНК, близкородственная «Alu-семейству» последовательностей ДНК с большим числом повторов. Блокирование трансляции не снимается до тех пор, пока комплекс СРЧ-лидерная последовательность – рибосома не свяжется с так называемым «отстригающим» белком (рецептором для СРЧ) эндоплазматического ретикулума. В этот момент начинается котрансляционное встраивание в эндоплазматический ретикулум. В процессе элонгации оставшейся части белка он перемещается через липидный бислой, поскольку рибосома остается присоединенной к эндоплазматическому ретикулуму. Таким образом образуется шероховатый (усеянный рибосомами) эндоплазматический ретикулум. Рибосомы остаются прикрепленными к эндоплазматическому ретикулуму втечении всего времени синтеза мембранного белка и освобождаются и диссоциируют на соответствующие субъединицы только после его завершения. Когда ранее синтезированная часть белка выходит в просвет эндоплазматического ретикулума, отщепляется лидерная последовательность, и присоединяются углеводы.
Интегральные мембранные белки не пересекают мембрану целиком; по-видимому, этому препятствует гидрофильная якорная последовательность на С-конце. Секретируемые же белки проходят сквозь мембранный бислой полностью и высвобождаются в просвет эндоплазматического ретикулума. К моменту их поступления внутрь везикулы углеводные остатки уже оказываются связанными с ними. Впоследствии секретируемые белки обнаруживаются в просвете аппарата Гольджи, где происходит модификация их углеводных цепочек, а затем они перемещаются к специфическим внутриклеточным органеллам или клеточным мембранам либо секретируются. Некоторые белки пересекают одну мембрану, а затем заякориваются в другой, соседней мембране, например внутренней мембране митохондрий.
2.2 Мембранная триггерная гипотеза
В этой гипотезе особое значение придается роли лидерной последовательности в изменении третичной структуры самого белка. Согласно этой гипотезе, лидерная последовательность индуцирует такую упаковку обычно гидрофобного интегрального белка, что последний может оставаться солюбилизированным в водной среде цитоплазмы, где он синтезирован. Мембранный липидный бислой является как бы триггером по отношению к третичной структуре белка – последний переходит в такую конформацию, которая обеспечивает его предпочтительное включение в бислой. Таким образом, белок претерпевает некий переход и сам встраивается в мембрану таким способом, чтобы установить необходимую поперечную асимметрию. Сразу после встраивания белка или его интеграции лидерная последовательность отщепляется. Триггерная гипотеза не предполагает специфического взаимодействия между рибосомой и мембраной, но это еще не означает, что синтез белка не может происходить на мембранах. Возможно, в одной и той же клетке действуют оба механизма.
3. Полипептидные сигналы, отвечающие за сортировку белков и
встраивание их в мембраны
Об аппарате и механизме переноса почти ничего неизвестно, немного больше известно о сигнальных последовательностях, присутствующих в полипептидах и направляющих каждый белок в нужное место. Успехов в этой области удалось достичь благодаря использованию техники рекомбинантных ДНК. С ее помощью были сконструированы гибридные полипептиды, в которые была включена тестируемая аминокислотная последовательность, принадлежащая другому белку. Таким образом можно было изучать влияние предполагаемой сигнальной последовательности на локализацию «белка-пассажира». Преимущества такого подхода удается использовать только в том случае, если вся информация, определяющая локализацию конечного продукта, заключена в первичной последовательности сигнала и если «белок-пассажир» является нейтральным участником процесса и, что существенно, подчиняется сигналу. Это условие выполняется во многих случаях, но известны и такие примеры, когда эффективность переноса или даже конечная локализация зависят от «белка-пассажира». Если «белок-пассажир» находится в конформации, не способной к переносу, то может происходить блокирование переноса химерного белка. Кроме того, функция некоторых сигнальных последовательностей зависит от их локализации в полипептиде или от взаимодействия с другими участками полипептидной цепи.
Похожие работы
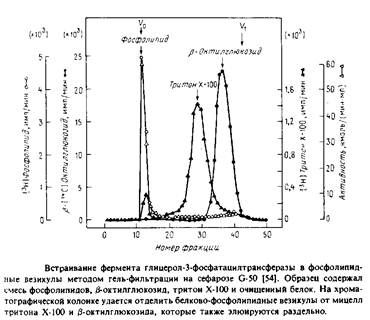
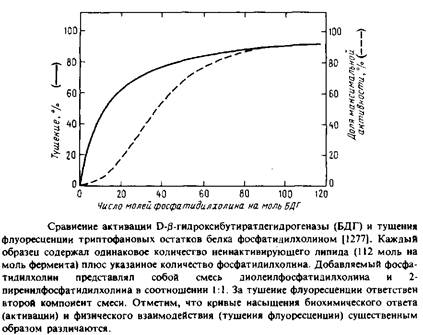
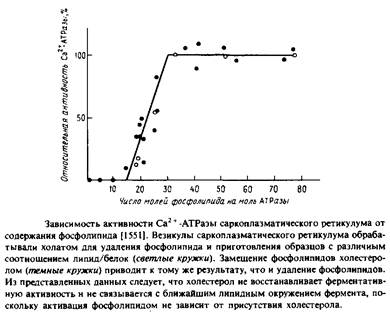
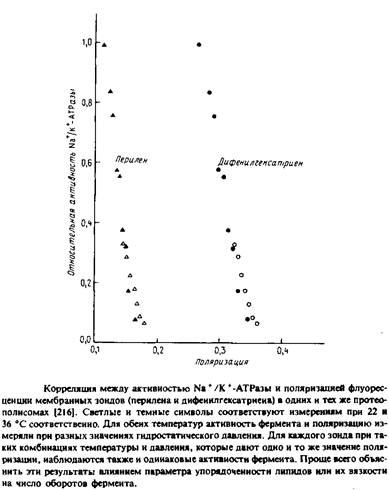
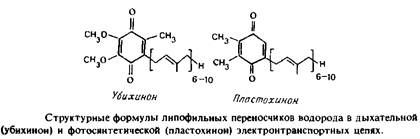
... . Все четыре системы являются основными белковыми компонентами мембран, в которых они локализованы, и могут служить иллюстрацией различных уровней организации мультиферментных комплексов в мембранной энзимологии. Первые две системы катализируют анаболические и катаболические реакции, протекающие в присутствии молекулярного кислорода и обычно липофильных мембраносвязанных субстратов. Терминальные ...
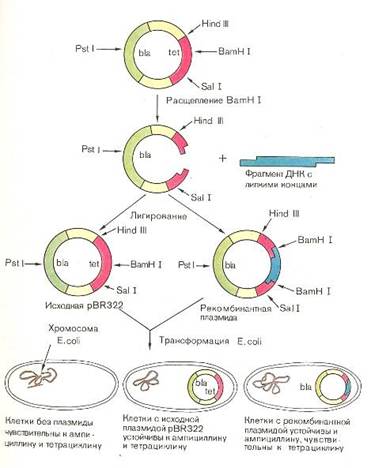
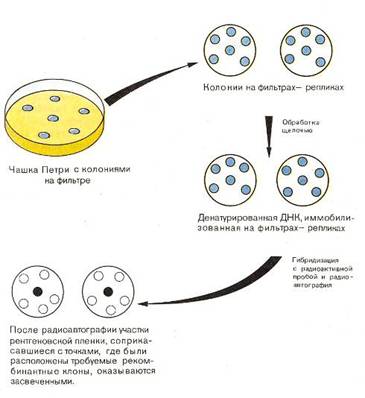
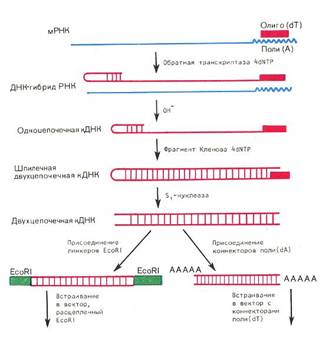
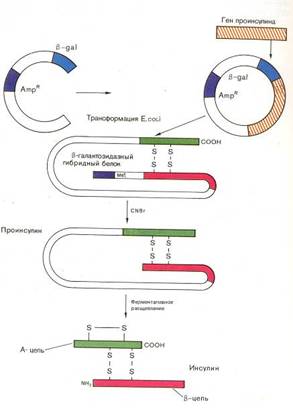
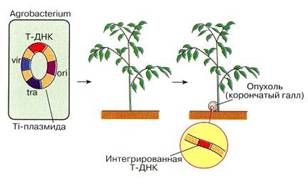
... инженерию. Необходимо отметить, что если базовый стандарт по химии не предусматривает изучение вопросов биотехнологии, то таковой по биологии содержит наиболее общие её аспекты: достижения генной инженерии и перспективы биотехнологии. 2.2 Межпредметные связи по изучению аспектов биотехнологии в средней школе По программе Р.Г. Ивановой и Л.А. Цветкова в 10 классе предусмотрено изучение темы ...
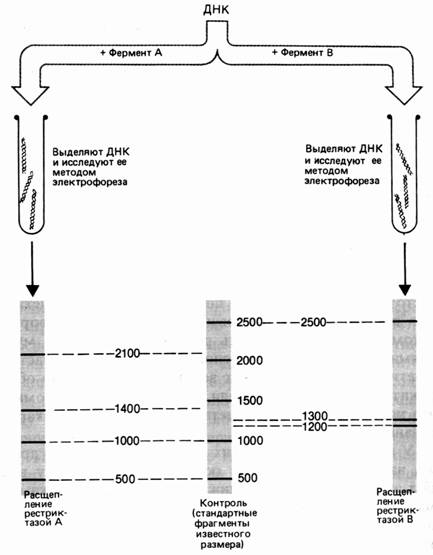
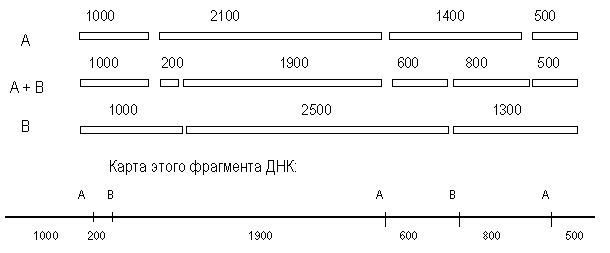
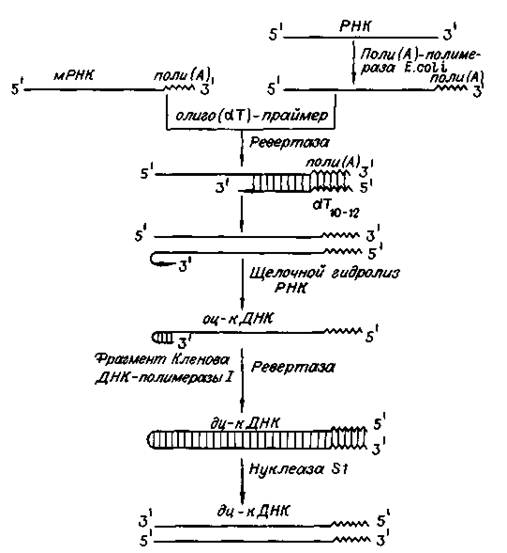
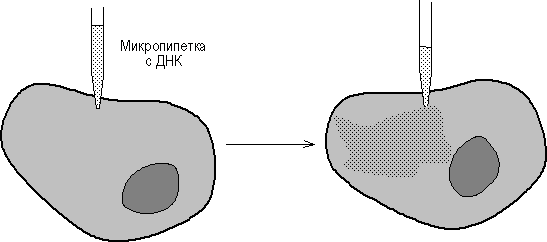
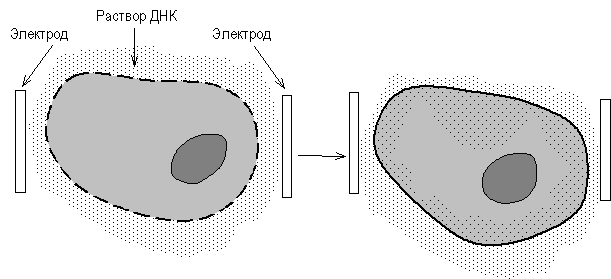
... векторов для экспрессии. 2.2 Способы прямого введения гена в клетку Прямое введение гена в клетку осуществляют несколькими способами: 1. Трансфекция 2. Микроинъекция 3. Электропорация 4. Метод «мини-клеток» 5. Упаковка в липосомы 6. Электронная пушка При трансфекции ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, ...
0 комментариев