Навигация
Методы исследования переноса белков через мембраны
36107
знаков
0
таблиц
4
изображения
1. Методы исследования переноса белков через мембраны
Наиболее детально изучены бесклеточные системы, в которых гораздо легче количественно исследовать процессы переноса и протеолитического процессинга белков. Во всех этих системах используются мембранные везикулы или препараты органелл, у которых поверхность, обращенная в цитоплазму, «смотрит» наружу, поскольку перенос белков осуществляется из цитоплазмы. Этому условию удовлетворяют микросомы, полученные из эндоплазматического ретикулума секретирующих клеток, митохондрий и хлоропластов. Вывернутые везикулы можно получить из клеток Е. coli, они представляют собой удобный объект для изучения переноса белков в бесклеточной системе.
Полипептид-предшественник, находящийся во внешней среде, при соответствующих условиях будет переноситься внутрь пузырька или, по крайней мере, через мембрану пузырька или органеллы. За этим процессом обычно следят, добавляя протеазы во внешнюю среду. Степень защиты от протеолиза является мерой количества полипептида, транспортированного внутрь везикулы или органеллы. За ходом протеолитического процесса, осуществляемого сигнальной пептидазой, следят с помощью электрофореза в полиакриламидном геле в присутствии ДСН. Белки, встроившиеся в мембрану, можно идентифицировать с помощью щелочной экстракции, при этом предполагается, что белки, которые связаны с поверхностью мембран, при такой обработке удаляются. Однако так бывает не всегда, поэтому результаты, полученные с помощью щелочной экстракции, необходимо интерпретировать с осторожностью.
В таких бесклеточных системах можно изучать биохимические условия переноса белков и идентифицировать необходимые растворимые компоненты. Кроме того, при этом можно варьировать природу переносимого полипептидного «субстрата».
При изучении бесклеточных систем были получены весьма важные данные об условиях, необходимых для переноса белков.
1. Посттрансляционный и котрансляционный перенос. Принято считать, что во всех исследованных системах перенос в мембраны или через мембраны может осуществляться независимо от трансляции. Убедительные данные на этот счет были получены для процесса переноса белков в хлоропластах и митохондриях, а также для переноса через бактериальную мембрану. Долгое время считалось, что перенос белков в эндоплазматический ретикулум или через мембраны эндоплазматического ретикулума всегда осуществляется параллельно трансляции, однако было четко показано, что такая параллельность не обязательна. Также важным считается то, что энергия, необходимая для переноса, не исходит от рибосомного биосинтетического аппарата.
2. Энергетические требования к переносу. Как правило, перенос белков в мембраны или через них энергозависим. Необходимым условием переноса как для прокариотических, так и для эукариотических систем является гидролиз АТР (или другого нуклеозидтрифосфата). Это было показано для следующих процессов: а) переноса белков в строму хлоропластов; б) транспорта белков в митохондриальный матрикс, внутреннюю и наружную мембраны; в) переноса белков через эндоплазматический ретикулум дрожжей и посттрансляционного встраивания мембранных белков в эндоплазматический ретикулум млекопитающих; г) переноса белков через цитоплазматическую мембрану Е.coli.
Еще одним независимым условием переноса белков в матрикс митохондрий и во внутреннюю мембрану митохондрий является наличие на последней трансмембранного потенциала. Этот потенциал, очевидно, необходим на ранней стадии процесса, при связывании белка с митохондрией.
3. Способность предшественника к переносу. Имеются веские доводы в пользу того, что ключевую роль в успешном переносе белка играет его четвертичная структура. Скорее всего это связано с тем, что сигнальная последовательность(ти), узнаваемая аппаратом переноса, должна быть доступна для него. Следовательно, для осуществления переноса белок должен быть неплотно свернут или частично развернут. Кроме того, если белки переносятся через мембрану в вытянутой конформации, то аппарат переноса должен быть способен к их развертыванию во время самого процесса переноса. Если бы белки- предшественники обладали стабильной четвертичной структурой, то они с трудом развертывались бы и, следовательно, не были бы способны к переносу.
Транспорт белков осуществляется в развернутом виде. АТР необходим для разворачивания полипептида. Разворачивание происходит до переноса или параллельно ему. На то, что именно АТР необходим для этого процесса, говорит тот факт, что транспорт укороченных предшественников в отличие от транспорта полноразмерного белка может осуществляться в отсутствии АТР. Впервые эти данные были сделаны на основе изучения митохондриальной мембраны. Для предотвращения свертывания предшественника в нативную конформацию необходим какой-либо растворимый белковый кофактор. Так, был выделен в водорастворимой форме, сходный порином митохондрий, предшественник белка наружной мембраны Е.coli OmpA, который был не способен к эффективному переносу через плазматическую мембрану, если в цитозоле отсутствовал белок, называемый «триггер-фактором». Известно также, что для переноса белков через мембраны эндоплазматического ретикулума млекопитающих или в эндоплазматичекий ретикулум необходим растворимый кофактор, а именно – сигнал-распознающая частица (СРЧ). Возможно, роль этого фактора состоит в предотвращении сворачивания предшественника полипептида.
Похожие работы
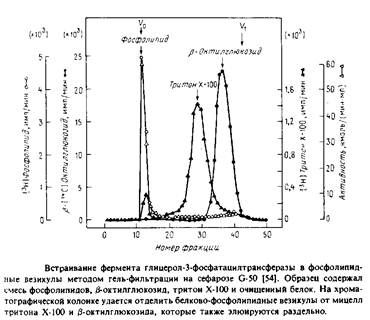
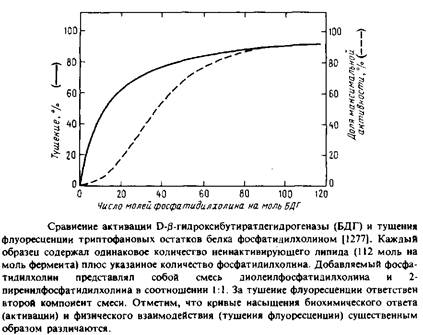
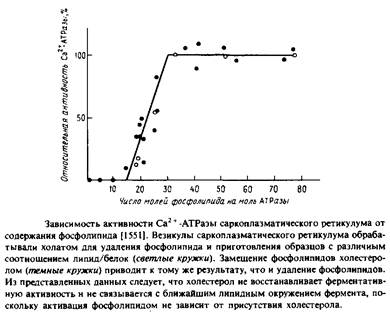
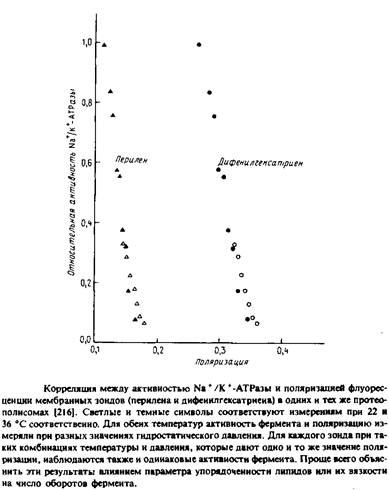
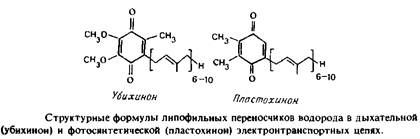
... . Все четыре системы являются основными белковыми компонентами мембран, в которых они локализованы, и могут служить иллюстрацией различных уровней организации мультиферментных комплексов в мембранной энзимологии. Первые две системы катализируют анаболические и катаболические реакции, протекающие в присутствии молекулярного кислорода и обычно липофильных мембраносвязанных субстратов. Терминальные ...
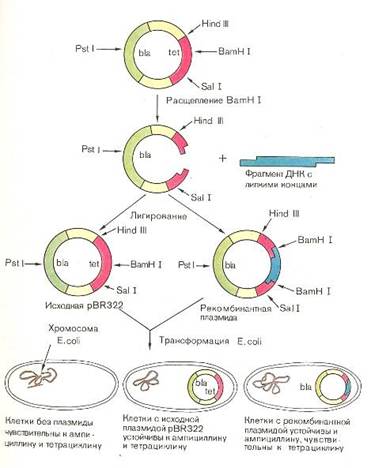
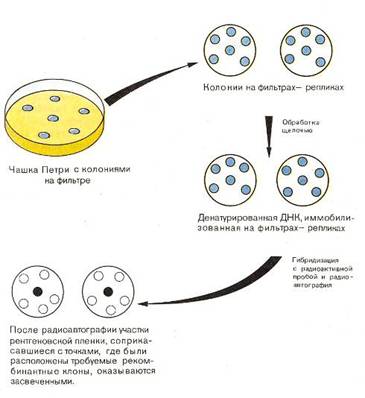
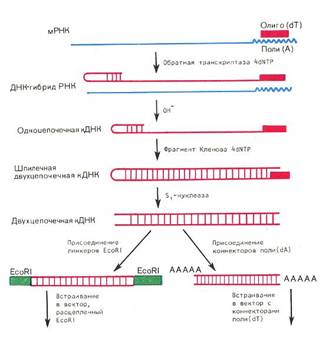
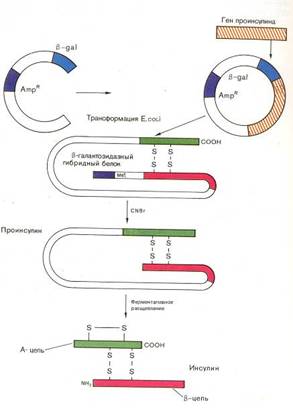
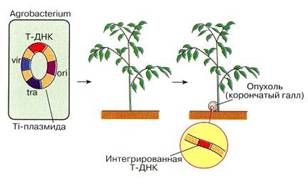
... инженерию. Необходимо отметить, что если базовый стандарт по химии не предусматривает изучение вопросов биотехнологии, то таковой по биологии содержит наиболее общие её аспекты: достижения генной инженерии и перспективы биотехнологии. 2.2 Межпредметные связи по изучению аспектов биотехнологии в средней школе По программе Р.Г. Ивановой и Л.А. Цветкова в 10 классе предусмотрено изучение темы ...
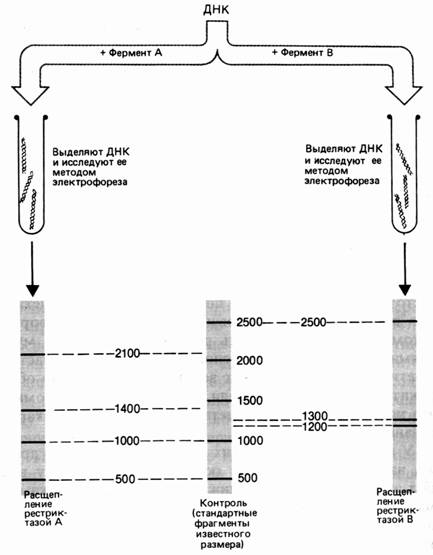
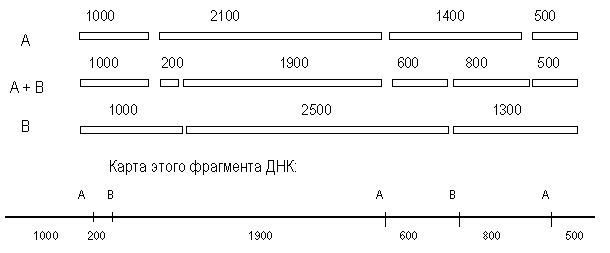
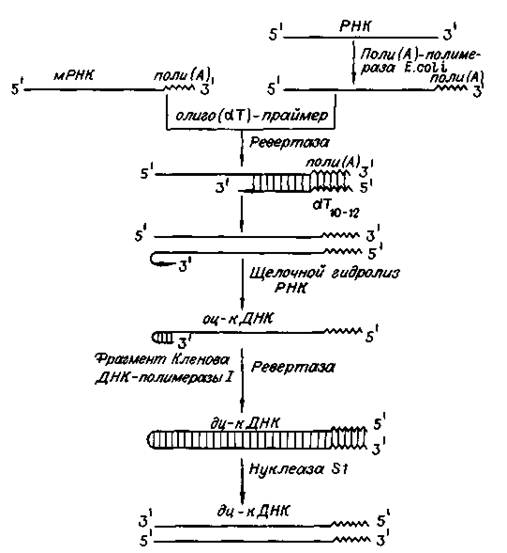
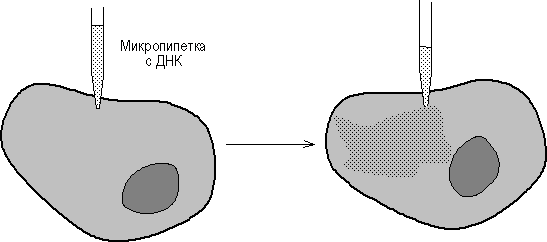
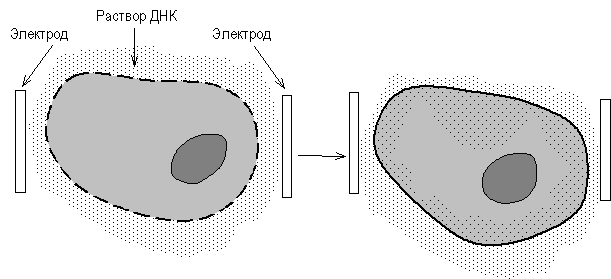
... векторов для экспрессии. 2.2 Способы прямого введения гена в клетку Прямое введение гена в клетку осуществляют несколькими способами: 1. Трансфекция 2. Микроинъекция 3. Электропорация 4. Метод «мини-клеток» 5. Упаковка в липосомы 6. Электронная пушка При трансфекции ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, ...
0 комментариев