Навигация
Генетична система мітохондрій
46998
знаков
1
таблица
2
изображения
1.2 Генетична система мітохондрій
Мітохондрія має власну генетичну систему - мітохондріальну ДНК і власний білоксинтезуючий апарат. Діленню (репродукції) мітохондрій передує реплікація мітохондріальної ДНК. Синтез ДНК мітохондрій при діленні клітин відбувається незалежно від реплікації ядерної ДНК.
У більшості об'єктів ДНК мітохондрій являє собою дволанцюгову ковалентно замкнуту структуру. В одній мітохондрії може бути від однієї до 20 000 кільцевих молекул ДНК, молекулярна маса яких варіює від (9-10) • 106 у хребетних до (3-4) • 107 у вищих рослині і грибів. Розміри кільцевих молекул ДНК у більшості мітохондріальних геномів варіюють від 16-20 (у хребетних) до 570 кілобаз у вищих рослин (кукурудза), а лінійна довжина молекули складає 5 мкм у дріжджів і 30 мкм у вищих рослин.
У мітохондріях тваринних клітин знаходиться кілька молекул ДНК, практично не помітних на звичайних електронно-мікроскопічних препаратах. У клітинах рослин їх, як правило, більше, так що при ультраструктурних дослідженнях мітохондріальна ДНК може бути виявлена у виді тонких фібрил. І, нарешті, у мітохондріях деяких кінетопластид (наприклад, трипаносомових) безліч кільцевих молекул ДНК поєднуються у велику електронно-щільну структуру - кінетопласт. У єдиній гігантській мітохондрії кінетопластид виявляється цікава модифікація мітохондріального геному: генетичний апарат тут представлений великою кількістю кільцевих молекул двох типів - так званих максі-кілець (вони гомологічні кільцевим ДНК мітохондрій інших об'єктів) і мінікілець - вони майже не містять генетичної інформації і їхнє функціональне значення стало прояснятися лише останнім часом. У рідких випадках, наприклад у водорості хламідомонади і інфузорій парамецій, мітохондріальний геном може складатися з лінійних молекул ДНК.
Про надмолекулярне упакування мітохондріальної ДНК відомо небагато. Очевидно, фібрили мітохондріальної ДНК за способом упакування нагадують хромосому прокаріот. У деяких об'єктів виявлені основні білки невеликої молекулярної маси, начебто зв'язані з мітохондріальною ДНК. Мітохондріальна ДНК відрізняється від ядерної за складом основ, плавучій щільності і ряду інших фізико-хімічних властивостей.
Значних успіхів досягла генетика мітохондрій. Численні роботи проводяться на мітохондріальних мутантах дріжджів (мутації дихального ланцюга, які не ведуть до загибелі кліток, тому що вони переходять на бродильний тип обміну). При дослідженнях клітин, які культивувалися in vitro вдалося одержати численні мітохондріальні мутанти в птахів, ссавців (у тому числі і людини); були отримані навіть клітини з мітохондріями без власної ДНК.
Дослідження в області мітохондріальної генетики з використанням мутантних клітин дозволили встановити, що ДНК мітохондрій властиві всі генетичні функції: рекомбінація, репарація і т.д. Було показано, що мітохондрії характеризуються так називаною полярністю, тобто перевагою в мітохондрій дочірніх клітин рекомбінантів одного типу: успадковування мітохондріального геному у тваринних і рослинних клітин в основному іде по материнській лінії. Правда, ці дані були отримані на обмеженому числі об'єктів. Останнім часом були виявлені факти спадкування обох типів батьківських мітохондріальних геномів у тваринних клітин і спадкування тільки по батьківській лінії в рослинних клітинах (секвойя). Для позначення цього явища - наявності в індивіда більше одного класу мітохондріального генома (або генома хлоропластів) запропонований спеціальний термін - гетероплазмія.
Дослідження в області мітохондріальної генетики дозволили картувати мітохондріальні геноми в різних об'єктів. Зокрема, є повна характеристика геномів мітохондрій клітин людини і дріжджів; про інші організми існують уривчасті відомості.
Обсяг інформації, закодованої в геномі мітохондрій, відносно невеликий; мітохондріальний геном, як правило, містить матриці для синтезу власних тРНК і високомолекулярних рРНК і деяких субодиниць ферментів дихального ланцюга й АТФ-синтазного комплексу.
Автономність білоксинтезуючої системи мітохондрій значно менша, ніж здається на перший погляд. Білкові фактори трансляції мітохондріальних РНК кодуються не ДНК мітохондрій, а ядерною ДНК, тобто процес трансляції в мітохондріях не може здійснюватися незалежно від ядерного геному. Крім того, у ДНК ядра є інформація, необхідна для синтезу факторів реплікації мітохондріальної ДНК, РНК-полімераз, що здійснюють транскрипцію ДНК мітохондрій, білків, які входять до складу 70 S рибосом (за винятком одного білка малої субодиниці) і більшої частини компонентів електроннотранспортного ланцюга й АТФ-синтазного комплексу.
В даний час велика увага приділяється вивченню механізмів транслокації білків через мітохондріальні мембрани. Встановлено, що численні білки дихального ланцюга, АТФ-синтазного комплексу і матриксу проходять у мітохондрії, очевидно, у ділянках тісного контакту зовнішньої і внутрішньої мембран - так називаних контактних сайтах. Тут знаходиться спеціальний білок-рецептор молекулярною масою 45 кДа, і утворюються специфічні канали. Процес цей вимагає енергії АТФ.
Більшість мітохондріальних мембранних ліпідів синтезується на ЕПС і вбудовується в мембрану мітохондрій. Ліпіди надходять до зовнішньої мембрани мітохондрій і попадають у внутрішню мембрану через контактні сайти. У мітохондріальних мембранах відбувається перетворення екзогенного фосфатидилсерину у фосфатидилетаноламін і утворення з «імпортованих» ліпідів кардіоліпіну - специфічного ліпіду внутрішньої мембрани мітохондрій.
Дослідження геному і білоксинтезуючого апарату мітохондрій дозволили прийти до висновку, що їм властиві деякі специфічні риси. Так, у мітохондріях має істотні особливості їх генетичний код. Універсальний термінінуючий кодон UGA у мітохондріях ссавців, дрозофіл і дріжджів кодує триптофан, а ізолейциновий кодон AUA - метіонін. Більш того, мітохондріальний генетичний код відрізняється в різних організмів. Так, лейциновий кодон GUA у мітохондріях ссавців і дрозофіл також кодує лейцин, а в мітохондріях дріжджів - треонін. Кодони AGA і AGG, які несуть в універсальному коді інформацію для синтезу аргініну, функціонують як термінуючі кодони в мітохондріях ссавців, кодують серин в мітохондріях дрозофіл і аргінін у мітохондріях дріжджів. У вищих рослин мітохондріальний генетичний код збігається з ядерним. Для зчитування всіх кодонів у білоксинтезуючому апараті мітохондрій досить всього 22-24 тРНК на відміну від цитозолю (де є принаймні 31 тРНК) і хлоропластів (30 тРНК); трансляція в мітохондріях відбувається з меншою точністю; мітохондріальні рибосоми також мають деякі особливості.
Геном мітохондрій людини відносять до так називаного „ощадливого” типу. Основна інформація закодована у важкому ланцюзі (Н-ланцюг) ДНК (у ній локалізуються гени двох високомолекулярних рРНК, 14 тРНК і 12 генів, які кодують іРНК для білків внутрішньої мембрани мітохондрій). У легкому ланцюзі (L-ланцюг) ДНК зберігається інформація тільки про 8 тРНК і одну іРНК для єдиного білка малої субодиниці мітохондріальних рибосом.
У мітохондріях рослин кодується низькомолекулярна РНК рибосом. У мітохондріальних геномах хламідомонади й інфузорії тетрахімени зберігається інформація не про всі тРНК: частина тРНК, які працюють у мітохондріях, закодована в геномі ядра. А в ядерному геномі трипаносомід представлена інформація про всі тРНК мітохондріальні білоксинтезуючі системи. Істотні розходження спостерігаються між різними об'єктами й у кодуванні білків дихального ланцюга й АТФ-синтазного комплексу. Таким чином, на тлі відносної сталості інформаційної ємності мітохондріального генома виявляється і значна еволюційна пластичність цієї ознаки.
Похожие работы
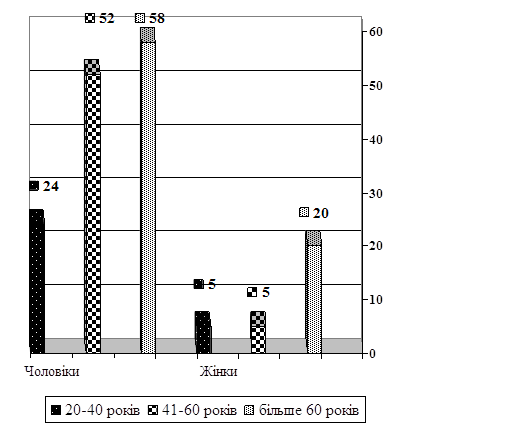
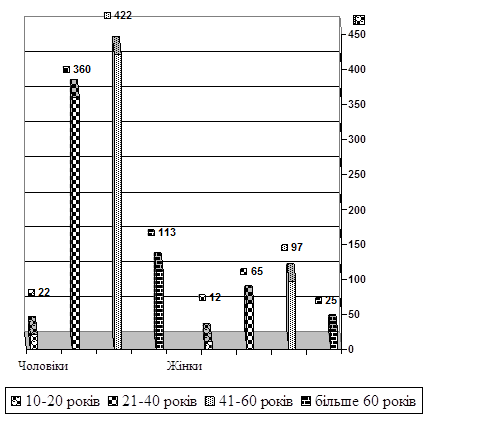
... 10.09 2007.- Бюл. №14. (особисто здобувачем виконані збір матеріалу та морфологічні дослідження). АНОТАЦІЯ Андре Мігель Ошкар Панзо. Можливість судово-медичної діагностики раптової серцевої смерті за результатами лабораторних досліджень. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.01.25 – судова медицина. – Луганський державний медичний уні ...
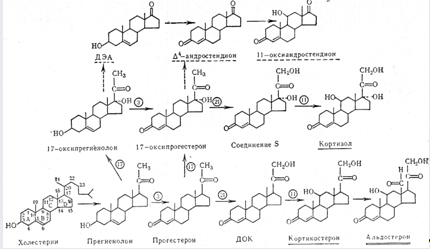

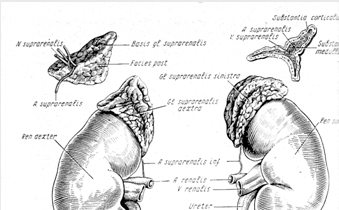

... контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою. Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо. Кіркова і ...
... . – Симферополь, 2008. – С. 35. (Здобувачем проведено набір матеріалу, аналіз даних, технічна підготовка до друку). АНОТАЦІЯ Шимкус Т.С. Вікові особливості будови легень під впливом гравітаційних перевантажень та при застосуванні різних засобів їх корекції (анатомо-експериментальне дослідження). - Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю ...
... ільш виражений у дівчаток від 10 до 11 років і у хлопчиків від 10 до 12 років (Таблиця 2). Показники функціонального стану повітряносних шляхів і легеневої тканини змінюються в тісному зв’язку із змінами антропометричних характеристик організму дітей на даному етапі онтогенезу. Особливо тісний зв’язок виявлений з довжиною і площею поверхні тіла. В перехідному періоді з «другого дитинства» до під ...
0 комментариев